Plenary04: Application of modern endocrine methods to conservation biology
Susumu Ishii
Department of Biology, School of Education, Waseda University, Tokyo 169-8050, Japan, fax 81 3 3207 9694, e-mail susumu@mn.waseda.ac.jp
Ishii, S. 1999. Application of modern endocrine methods to conservation biology. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban. Ostrich 70 (1): 33–38.
I report here on novel endocrinological methods originally developed for the artificial breeding of the Japanese Crested Ibis, Nipponia nippon. According to the most recent information, the only wild population in the world is one of about 60 individuals in Yang Xian, China. Captive populations of 54 and 17 individuals are located in Yang Xian and at Beijing Zoo, respectively and a single female of this species survives in Japan. Females of many birds stop breeding when they are brought to captivity. Hormone administration is considered to be effective in stimulating the breeding activity of such females. If the ovary of a captive female contains well-developed ova but does not ovulate, the problem is not serious. Ovulation can be induced by injection of gonadotropic hormone. However, if the ovary is regressed and contains no developed ova, the problem is serious. Until recently, no avian endocrinologist had been able to induce ovulation in hens with a regressed ovary. We developed a non-invasive method to assess the ovarian and testicular condition of birds by analysing gonadal sex steroid hormones in droppings. By using this method, we can decide whether the ovary contains developed ova or not and whether a male is sexually active or not. To reactivate a regressed ovary, we employed gonadotropin administration by means of the osmotic pump implantation in female Japanese Quail Coturnix japonica kept under short-days and then injected with the gonadotropin preparation once a day for several days. About 50% of the females laid an egg or eggs in response to the injection. Seven of the eggs were artificially incubated and two of them hatched. One of the two was revealed to be a male and fertile. The faecal sex steroid assays have been successfully applied to males of the Japanese Crested Ibis and Kakapo Strigops habroptilus in New Zealand. Although too late to apply the hormone administration method to the Japanese Crested Ibis for breeding, application of the method to the Kakapo is now being examined in New Zealand.
INTRODUCTION
Captive breeding is one of the methods that can be used to save endangered avian species. The final goal of a captive breeding program must be to ensure recovery of the wild population by releasing captively bred individuals to their original, or newly established habitat. The Toki, or Japanese Crested Ibis Nipponia nippon (order Ciconiformes, family Threskiornithidae), is a critically endangered species in Japan. Since 1982, attempts at captive breeding of this species have been conducted in Japan but, unfortunately, this program is now going to be closed with no substantial result for the Toki itself. During the course of the attempts, however, we developed some endocrine methods that are considered to be useful in conservation biology. In the present paper, we briefly describe the past and current status of Japanese and Chinese populations of the Toki, and introduce the methods we developed. In addition, we also report on our program to preserve cells and tissues of the Toki.
Toki in Japan
According to a beautiful monograph by Yamashina and Nakanishi (1983), written in Japanese, Toki were distributed only in the north-eastern half of Japan and were uncommon birds in the 17th and early 18th centuries. Later, they expanded their distribution and became rather common birds in the late 18th and 19th centuries. Literature in the 18th century indicated that Toki were observed in part of Hokkaido, various parts of Honshu Island, some parts of Kyushu, and small islands around Honshu and Kyushu (Fig. 1). In the 1750s, Toki were introduced artificially to Shikoku, another big island of Japan, and became common at least locally. Thus, the Toki was distributed all over Japan except parts of Hokkaido, and was commonly observed in these areas in the late 19th century. Yamashina and Nakanishi (1983) reported that the expansion of the distribution of this species to western Japan was mainly due to artificial introductions. In the early 20th century, the Toki suddenly started to disappear everywhere in Japan, and it was once believed that the Toki was extinct by the 1920s. A small number of Toki were rediscovered on Noto Peninsula and Sado Island in 1929 and 1931, respectively. However, these two populations tended to decrease their sizes year by year and the Noto population had completely disappeared by 1970 (Fig. 2).
The last five individuals of the Sado population were captured in 1981 and housed in the Toki Preservation Centre on Sado Island together with a female named ‘Kin’ that was captured in 1968. The captive breeding program thus started with these six birds consisting of a male and five females. Soon after, two females died presumably due to staphylococcal infection. The program was then conducted using two females and the only male. One of the remaining three females was excluded from the program because of a physical problem with one of her legs. Recently, a small number of Chinese Toki, kindly loaned by the Chinese Government, also joined the program. However, all breeding trials using these birds have failed and most of the birds died one by one (Fig. 2). Only one female, 31 years old, survives now.
Toki in Korea and eastern Siberia
Toki were observed in a number of places all over Korea and in an area along Usurri River in eastern Siberia (Fig. 3) until the early 20th century. However, according to the monograph by Yamashina and Nakanishi (1986), both populations declined in this century and the last individual in Korea was observed in 1979 and in east Siberia in 1981. Toki disappeared approximately at the same time in both areas and also in Japan.
Toki in China
The Toki was widely distributed in the northern half of China, Taiwan and Hainang Island in the 19th to early 20th centuries (Fig. 3). In China as well as in other Asian countries, the Toki suddenly started to disappear and it was also thought that no wild population remained in China. A team of scientists from the Zoology Institute, Academia Sinica, Beijing, started a survey of the Toki in 1978. After surveying more than ten provinces, they at last discovered two adult pairs and three young individuals in two nests at small villages near Yang Xian in Shaanxi Province in 1981 (Liu & Katsura 1992). Since that time, the Chinese Government has made efforts to protect this small population in Yang Xian. At the same time, they captured young individuals fallen from nests and attempted captive breeding. According to the most recent information, the sizes of the wild and captive populations in Yang Xian are about 60 and 54, respectively, and there are additional 17 captive individuals breeding at Beijing Zoo (personal communication from Dr Kaneko of Toki Preservation Centre who visited Yang Xian in June 1998). Thus, both the wild and captive populations in China are increasing in size.
ENDOCRINE METHODS FOR ARTIFICIAL BREEDING OF BIRDS
Two years after the start of the captive breeding program of the Toki sponsored by the Niigata Prefecture Government and the Environment Agency of Japan, the author was invited to join the program and asked to examine the possibility of employing hormonal treatment to stimulate ovarian activities in the Toki. At that time, there remained only two females; one was old and another was unhealthy, and a male. It seemed to be too late to apply the hormonal replacement therapy in Toki. However, the Environment Agency and Ministry of Culture, Science and Education, Japan and Niigata Prefecture supported us to develop endocrine methods for the artificial breeding of birds with a hope of applying the methods to other endangered birds. Recently, we also started a program to preserve cells, tissues and nucleic acids of Toki for future scientific use under the sponsorship of the Environment Agency.
Non-invasive assessment of gonadal endocrine activities by faecal sex steroid analysis
When a female bird does not ovulate, we can assume that there are two possibilities for the condition of the ovary. One is that the ovary contains fully-grown follicles but no ovulation occurs. The other is that the ovary is regressed completely and contains only small immature follicles. In the former case, it is easy to induce ovulation. As endocrinology textbooks teach us, an injection of luteinizing hormone that mimics the luteinizing hormone surge is effective at inducing ovulation in such a female. In the latter case, we have to stimulate the ovary first with follicle-stimulating hormone in order to stimulate ovarian follicles to develop and then inject luteinizing hormone to induce ovulation. Thus, the hormone treatment method will differ depending on the conditions of the ovarian follicles.
To establish the condition of the ovary in a female bird, it is best to directly observe the ovary during laparotomy. However, this is not the preferred method, of course, for captive individuals of precious rare birds. The second best method is the measurement of concentrations of follicle-stimulating hormone and luteinizing hormone or ovarian sex steroid hormones in blood plasma. Follicle-stimulating hormone stimulates ovarian follicles to grow and luteinizing hormone acts on the most developed ovarian follicle causing ovulation. The former hormone also stimulates the ovary to secrete oestrogen (mainly oestradiol 17beta) and the latter hormone regulates progestin release (progesterone in birds and mammals). We have some results showing levels of luteinizing hormone and follicle-stimulating hormone in blood plasma of four captive individuals of the Toki (Fig. 4). We used blood collected in 1982 from three females and a male. The hormone level was highest in Shiro (or White), a female that almost laid an egg but died, and second highest in a male Midori (or Green) that was mated with Shiro. The hormone level was low in the unhealthy female, Ao (or Blue) and lowest in the oldest female (Kin or Gold). In this study, we could confirm that the hormone level in plasma is a good index of the gonadal condition. However, frequent bleeding of birds is of course not the preferable method and a non-invasive method is necessary. Such a non-invasive method, the measurement of urinary sex steroid hormones, is frequently employed in mammals. In birds, as is well known, both urine and faeces are excreted together. Accordingly, we decided to extract sex steroid hormones from faeces (actually a mixture of faeces and urine) and measure their concentrations in faecal mass by conventional radioimmunoassays. Adult Japanese Quail, Coturnix japonica, were used as study birds for this work.
As shown in Fig. 5, testosterone concentrations in both plasma and faeces were drastically reduced by gonadectomy and also by a short-day treatment in males of Japanese Quail. The administration of oestradiol in different doses increased both plasma and faecal concentrations of the hormone in a dose dependent manner in ovariectomized female Japanese Quail (Fig. 6). Thus, we demonstrated that sex steroid hormone levels in plasma are faithfully reflected in concentrations of sex steroid hormones in faeces. We can therefore assess gonadal endocrine activities or gonadal condition by knowing sex steroid concentrations in faeces.
We attempted further to detect the occurrence of ovulation by measuring the concentration of progesterone in faeces collected successively through a period of a day in laying and non-laying hens of the Japanese Quail. It has been known that the LH surge, a transitory secretion of a large amount of luteinizing hormone, induces ovulation and a concomitant rise in the progesterone concentration in plasma. We expected that this change in the progesterone concentration would be reflected in the faecal progesterone concentration. We selected hens predicted to ovulate in the afternoon (laying quail) from a large number of Japanese Quail hens. At the same time, non-laying quail hens were also selected and used. We collected droppings of all these birds individually. The laying quail hens showed a clear peak in the mean faecal progesterone concentration around the time of expected ovulation that was estimated from the actual oviposition time (Fig. 7a). Non-laying quail hens showed no clear peak in the faecal progesterone concentration (Fig. 7b). If we can collect faeces successively from individual female bird during the breeding season, we may be able to estimate the time of ovulation using the faecal progesterone analysis. By using a modern automatic enzyme immunoassay system for sex steroid hormones, we may quantify progesterone levels in a sample within several minutes.
Application of the developed methods
We applied the faecal steroid hormone method to correlate faecal sex steroid levels with reproductive behaviour of Rock Ptarmigans Lagopus mutus of a wild population at Murodo Daira in Tateyama Mountains in Japan (Kikuchi & Ishii 1997). Sex steroid hormone concentrations in both male and female faeces changed in close association with changes in the reproductive behaviour of the birds. We also surveyed changes in testosterone, a testicular sex steroid hormone, in faeces of a male Japanese Crested Ibis from non-breeding months to the months just before the breeding season, and found that the testosterone concentration was elevated during December to January when the nuptial coloration was appearing (Fig. 8). Dr John Cockrem of Massey University, New Zealand employed this method to estimate the breeding period in Kakapo Strigops habroptilus and found that faecal testosterone concentrations rose rapidly during the breeding season (Cockrem 1997). Now we can use the faecal steroid hormone method as an excellent non-invasive tool to estimate the endocrine activity of the gonads in wild birds.
Artificial induction of ovulation by treatment with gonadotropic hormone
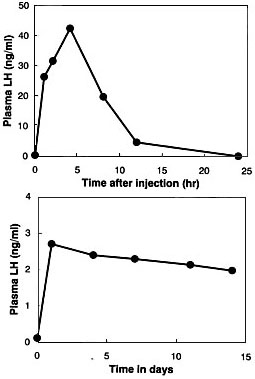
A number of poultry scientists have attempted to induce ovulation in completely sexually inactive hens of the domestic fowl or the Japanese Quail by injecting mammalian gonadotropic hormone preparations or avian pituitary extracts, but all of them failed to induce ovulation (Wakabayashi et al. 1992). We conducted a similar experiment using sexually inactive short-day females of the Japanese Quail and confirmed the previous results. Then, we studied changes in concentrations of luteinizing hormone and follicle-stimulation hormone after the injection of a chicken pituitary gland extract into a quail. The gonadotropin concentrations rose rapidly after the injection but they soon declined to extremely low levels (see Fig. 9a for luteinizing hormone). Finally, we used osmotic pumps (Alzet Company) to maintain high hormone levels. We loaded each pump with a glycoprotein fraction extracted from chicken pituitary glands and implanted the loaded pump into the body cavity of each female kept under a short-day photoperiod. Luteinizing hormone levels in plasma of the implanted female remained almost constant for about two weeks with this treatment (Fig. 9b). The ovaries of birds treated with hormones by this method contained fully grown follicles but, disappointingly, no ovulation occurred in the treated birds.
Then, we combined the two treatment methods, i.e. the chronic treatment by means of the osmotic pump and the short-term transitory treatment by injection. Short-day females were treated with chicken pituitary extract by means of Alzet osmotic pumps for about two weeks, and daily injections of the same extract were given during the last four days of the chronic treatment period (Wakabayashi et al. 1992). Four of the seven treated females laid seven eggs in total. All seven eggs were kept in an incubator and two male chicks hatched normally. After sexual maturation, one of them was examined for fertility by mating with a mature female. It was revealed that the male was indeed fertile.
We concluded that a combination of the two methods was effective to induce ovulation followed by oviposition in about 40% of treated female Japanese Quail. We further used commercial mammalian gonadotropin preparations (pregnant mare serum gonadotropin and human chorionic gonadotropin) instead of the chicken pituitary extract, and found that they were less effective than the chicken pituitary extract. It may be safe and most effective to use homologous gonadotropic hormones. This method can be a potential tool to breed captive birds artificially. At the same time, it is also useful to obtain mature ova and spermatozoa any time when we need them.
PRESERVATION OF TISSUES AND LIVING CELLS OF THE JAPANESE CRESTED IBIS
Preservation procedures
Recently, two males of the Japanese Crested Ibises died at the Sado Toki Preservation Centre successively in December of 1994 and April of 1995. The former was a Chinese Toki borrowed from the Chinese Government for breeding and the latter individual was one of five Toki captured on Sado Island in 1982 and subsequently reared at the Centre.
For the Chinese male, a part of the liver, a part of the kidney and both testicles were dissected out about 8 hours post-mortem and chilled on ice. At the same time, other parts of the liver, kidney, a part of the breast muscle and the whole pituitary gland were cut out and immediately frozen on dry ice. About two hours later, the frozen tissues were moved into a tank containing liquid nitrogen, and stored there. At the same time, the former tissues chilled on ice were minced with scissors into fine pieces and subjected to enzymatic digestion with a 0.2% trypsin solution for the liver and kidney and a 0.2% solution of collagenase for the testicles. Separated cells thus obtained were dispersed in a medium for cell culture and stored in the presence of an antifreeze reagent (20% dimethyl sulphonoxide or DMSO) in liquid nitrogen. Quantities of separated cells from these organs and their viabilities were determined soon after separation (Table 1 and Table 2).
For the Japanese male, Midori, we also applied similar procedures when it died in April of 1995. In this individual, living cells were separated from the liver, kidney, testis, lung and skin. They were cryo-preserved as mentioned above. Pieces of almost all Midori’s organs except the adrenal gland were stored in liquid nitrogen. Quantities and viabilities of the cryo-preserved cells are given in Table 2. Viability values in the cells of the organs of Midori were lower than the values in the Chinese individual. This difference may be because the viability was tested before cryo-preservation in the Chinese individual but after cryo-preservation in Midori, and also the cell separation procedure was started about eight hours after death in the former but ten hours after death in the latter bird.
In addition to these materials, skin cells of the last 6 individuals of the Japanese population are cryo-preserved in the Chromosome Research Station in Faculty of Science, Hokkaido University. Skin tissues of these birds were sent there for sexing by chromosome analysis soon after capture.
Why store cells and tissues?
Genetic studies
The genetic relationship between Chinese and Japanese populations has never been studied. The stored materials can be used for such a study. Genetic relations of different individuals in the Japanese population should be studied using the stored cells or tissues to provide information on genetic homogeneity or heterogeneity of the last six individuals of the Japanese population.
Cloning of gonadotropic hormone genes
For artificial induction of ovulation in Toki, it is preferred to use homologous gonadotropic hormones. Production of recombinant Toki gonadotropins must be conducted for this purpose. We extracted DNA from Midori’s cryo-preserved kidney tissue, and recently determined partial sequences of Toki gonadotropic hormone subunit genes using the extracted DNA as a template and authentic nucleotides with partial sequences of cDNA encoding gonadotropic hormone subunit molecules of the Japanese Quail as primers. We are now determining the full-length structures of the gonadotropic hormone subunit genes using a genomic library prepared from the preserved kidney tissue.
Production of Toki by using a related species and modern biotechnology
A technique for the implantation of primordial germ cells (embryonic cells which migrate to the gonad and become egg or sperm cells in future) to an embryo of a heterologous species is now being developed by some scientists (Ono & Yokoi 1994; Wentworth et al. 1980). A technique to implant a nucleus of a cell to another enucleated cell is now being commonly used. By combining these techniques, it may be possible to reproduce a Toki from the preserved cells. First, we collect primordial germ cells from an embryo of a species related to Toki, and replace their nuclei with those separated from preserved cells of an individual Toki. We may increase the number of these cells by culturing. Then, we implant the cultured primordial cells with somatic cell nuclei of Toki into an embryo of the related species. If the recipient is a male, some spermatozoa should have the donor nucleus and others the recipient nucleus. In this way, we can produce a female with a chimeric ovary. By mating such birds with chimeric gonads, we can obtain a fertilised egg of the donor type although this occurs by chance in a certain proportion.
It is also prerequisite to reconstruct an area good enough as a natural habitat of Toki before we complete this technique and reproduce Japanese Toki artificially. Only after the reconstruction of the natural habitat, can we produce a new field population of Toki in Japan.
ACKNOWLEDGEMENTS
Studies introduced in this paper were conducted in collaboration with Masaru Wada, Shuichi Wakabayashi, Motoshi Kikuchi, Nobuyuki Yamaguchi, Hidetsugu Sakai, Kaoru Kubokawa, Yoshiaki Izaike, Kazuhiro Kikuchi, Junko Noguchi and Yoshihiro Kubodera. The author thanks Tsunao Watanabe and Takako Sako, Environment Agency, Japan. Without their strong will and tireless efforts, the project for preservation of cells and tissues of the Toki could not be realised. Thanks are also due to Kohki Chikatsuji and Yoshinori Kaneko of the Toki Preservation Centre in Sado Island for supplying faecal samples of the Toki and information on the Toki and to Dr John Cockrem and Dr John Wingfield for their reviews of this manuscript.
REFERENCES
Cockrem, J.F. 1997. Contributions of comparative endocrinology to conservation biology. In: Advances in Comparative Endocrinology, S. Kawashima, S. & S. Kikuyama, S. (eds) Monduzzi Editore, Bologna, Italy, pp. 1695–1700.
Ishii, S., Wada, M., Wakabayashi, S., Sakai, H., Kubodera, Y., Yamaguchi, N. & Kikuchi, M. 1994. Endocrinological studies for artificial breeding of the Japanese Ibis, Nipponia nippon, an endangered avian species in Asia. J. Biosci. 19, 491–502.
Kikuchi, M. & Ishii, S.. 1997. Estimation of gonadal and adrenal activities in birds by analyzing faecal steroid hormones. In ‘Advances in Comparative Endocrinology’, S. Kawashima and S. Kikuyama eds., Monduzzi Editore, Bologna, Italy, pp. 1393–1398.
Liu, I. & Katsura, C. 1992. Tokiga ikite ita. Popurasha, Tokyo (in Japanese).
Ono, T. & Yokoi, R. 1994. Transfer of male and female primordial germ cells of quail into chick embyronic gonad of the same and opposite sexes. Proc. 5th World Congress on Genetics Applied to Livestock Production, 20, 104–107.
Yamashina, Y. & Nakanishi, G. 1983. Toki, Nipponia nippon. Kyoikusha, Tokyo (in Japanese).
Wakabayashi, S., Kikuchi, M., Wada, M., Sakai, H. & Ishii, S. 1992. Induction of ovarian growth and ovulation by administration of a chicken gonadotrophin preparation to Japanese Quail kept under a short-day regimen. Brit. Poult. Sci. 33, 847–858.
Wentworth, B.C., Tsai. H., Hallett, J.H., Gonzales, D.S. & Rajcicspasojevic, G. 1989. Manipulation of avian primordial germ cells and gonadal differentiation. Poult. Sci. 68, 999–1010.
Table 1. Quantities and viabilities of cells separated from organs of a Chinese male Toki Nipponia nippon which died in Japan. Viability was determined soon after cell separation and before cryo-preservation.
Table 2. Quantities and viabilities of cells separated from organs of a Japanese male Toki Nipponia nippon which died in Japan. Viability was determined after cryo-storage in liquid nitrogen for about one month.
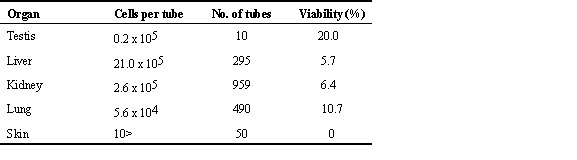
Fig. 1. Distribution of the Toki Nipponia nippon in Japan in the 17th to 18th centuries (based on Kuroda & Nakanishi 1983).
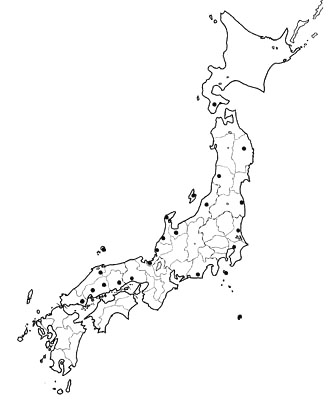
Fig. 2. Change in the sizes of the Noto Peninsula and Sado Island populations of the Toki Nipponia nippon after 1952 or 1953. The Sado Island population contained captive individuals. In January of 1981, the last five wild individuals of the Sado Island population were captured and moved to captivity.
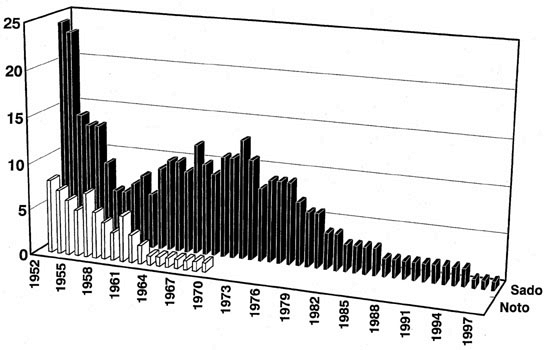
Fig. 3. Distribution of the Toki Nipponia nippon in Asia from the 19th to 20th centuries based on Kuroda and Nakanishi (1983) and Liu and Katsura, (1992). The location of the one existing wild population, Yang Xian, is circled.
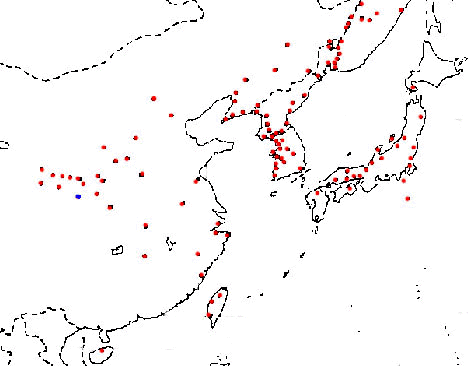
Fig. 4. Concentrations of luteinizing hormone (LH) and follicle-stimulating hormone (FSH) in the blood plasma of four captive Toki. Blood samples were collected in October 1982.
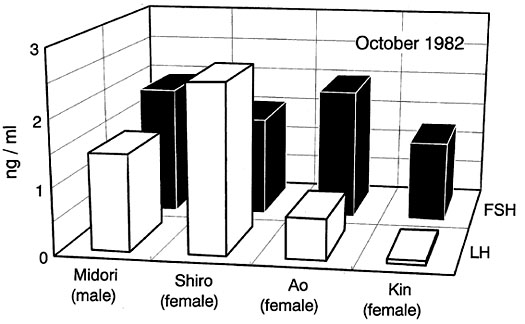
Fig. 5. Concentrations of testosterone in both blood plasma and faeces of sexually active adult Japanese Quail Coturnix japonica males kept at long-days (16L8D), sexually inactive males of the same age kept at short-days (8L16D) and castrated males of the same age.
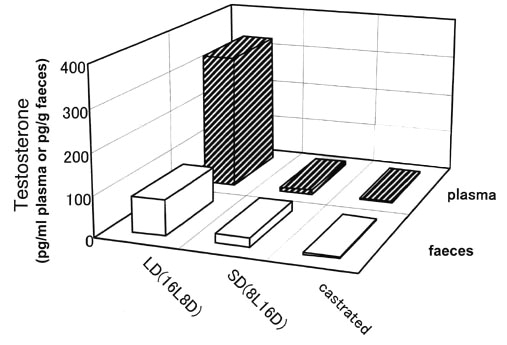
Fig. 6. Concentrations of oestradiol in both blood plasma and faeces of five ovariectomized female Japanese Quail Coturnix japonica implanted with different number of Silastic tubes containing a constant amount of oestradiol.
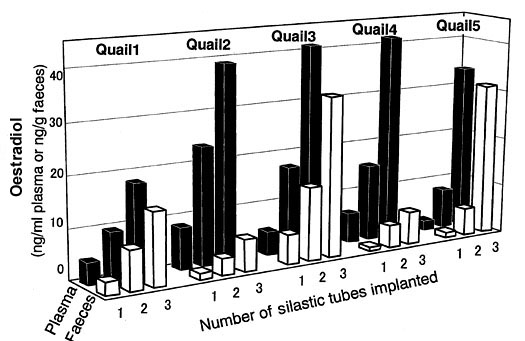
Fig. 7. Upper figure: Changes in the mean concentration of progesterone in faeces in seven laying Japanese Quail Coturnix japonica hens during a day. The period during which ovulation was estimated to occur is indicated with a thick line on the horizontal axis showing the time of the day. Lower figure: Changes in the mean concentration of progesterone in faeces in seven non-laying Japanese Quail hens during a day. The non-shaded and shaded portions of a bar under the horizontal axis show the light and dark periods, respectively.
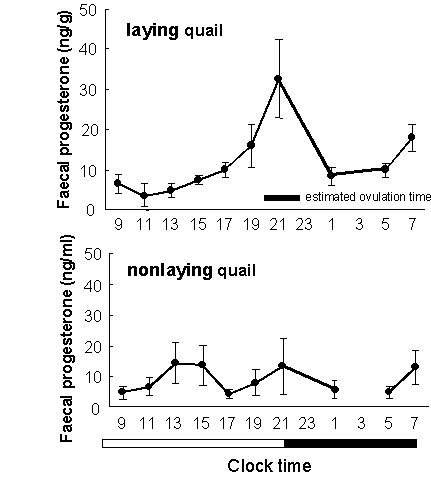
Fig. 8. Concentrations of testosterone in faeces of a male Toki, Midori. Samples were collected in July, August, September and a period from December to January. The concentration of testosterone in each period is expressed as the mean of four faecal samples collected on four successive days in each period. The vertical bar indicates the standard error of the mean.
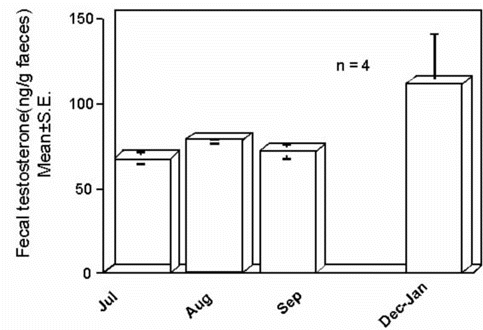
Fig. 9. Upper figure (9a): Changes in the concentration of luteinizing hormone in blood plasma in an ovariectomized Japanese Quail Coturnix japonica hen after an injection of a glycoprotein fraction extracted from chicken pituitary glands. Lower figure (9b): Changes in the concentration of luteinizing hormone in blood plasma in an ovariectomized Japanese Quail hen implanted with an osmotic pump loaded with a glycoprotein fraction extracted from chicken pituitary glands.