S18.4: Food supply and breeding activity of Skylarks in fields with different pesticide treatment
Niels Elmegaard1, Per Nørmark Andersen1, Peter Odderskær2 & Allan Prang2
1National Environmental Research Institute, Department of Terrestrial Ecology, PO Box 314, DK-8600 Silkeborg, Denmark, fax 45 8920 1414; e-mail
ne@dmu.dk; 2Department of Landscape Ecology, DK-8410 Rønde, Denmark Elmegaard, N., Andersen, P.N., Odderskær P. & Prang, A. 1999. Food supply and breeding activity of Skylarks in fields with different pesticide treatment. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1058-1069. Johannesburg: BirdLife South Africa.The risk of direct poisoning of birds in arable land due to pesticides is low and still declining in Denmark because the most toxic substances are disappearing from the market. The indirect effect of pesticide use on habitat quality may, however, be of great importance to species living in agricultural fields. In order to quantify the indirect effect of pesticide spraying on avian food supply, a study of Skylark Alauda arvensis L. breeding performance in fields subjected to different pesticide treatment was undertaken. Skylark reproduction, nestling food and food abundance were measured during four years in four spring barley fields, two subjected to full pesticide treatment and two treated only with fungicides and a seed dressing. Food abundance was reduced, nestling food was less diverse, nestling survival and number of nesting attempts lower in fields subjected to full pesticide treatment. Food abundance varied considerably between years and fields. Thus the importance of the indirect effect of pesticide applications on skylark breeding success also varied between years and fields. The relation between food abundance and breeding activity within a season was not simple. The interaction between food abundance, breeding success and pesticide use is complex in multi-brooded altricial species probably because several factors are governing the breeding effort invested by the parents in a particular brood.
INTRODUCTION
When cultivated fields are subjected to pesticide spraying the avifauna may be affected by the direct toxic effect of the chemicals and by the indirect effect derived from changes in habitat quality such as reduced food supply. The relative importance of the direct and the indirect effects on food availability depends on the specific toxicity of the compounds applied and the reduction of the food resource in relation to the food demand. Both factors are species specific. In Denmark the average acute toxicity towards birds of the pesticides on the market has declined during the last two decades due to changes in legislative procedures and the development and use of new compounds with lower bird toxicity (Clausen 1998).
In bird surveys from Denmark the decline in numbers of typical farmland species during the time period of rapid intensification of agriculture, has often partly been ascribed to the use of pesticides (Petersen 1996). In studies comparing the avifauna on conventional and organic farms (Braae et al. 1988) it also has been suggested that pesticides may play a role in the observed lower densities and species numbers on conventional farms. Studies of the Yellow Hammer Imberiza citrinella L. have demonstrated a higher reproductive success in organically managed fields (Petersen et al. 1995). The authors suggested that a poorer food abundance in conventional fields was the main reason for this difference and that pesticide use probably contributed to this reduction. The above mentioned studies did not provide any direct evidence of pesticide effects on food abundance or quality in surveyed fields. Furthermore, the influence of farming practices on the avifauna goes beyond pesticide use and includes factors such as crop type, fertilising, soil tillage, field size, boundary quality etc. The specific impact of pesticides can be difficult to extract from this multi-factorial complex of variables many of which are inter-dependent.
Many studies have illustrated the detrimental impact of insecticides on the arthropod fauna and a few have compared the arthropod fauna of organically and conventionally managed fields and found a significant difference in species composition (Moreby et al. 1994, Reddersen 1997). Also the weed flora differs between the two management systems (Hald and Reddersen 1990). Southwood and Cross (1969) drew attention to the importance of arthropod food abundance for chick survival, and to the weed - insects - Partridge food chain and how it is affected by herbicide spraying. At the Game Conservancy in the UK a number of studies have convincingly suggested a causal relationship between arthropods, gamebirds and pesticide use in arable land (Potts 1986). Rands (1986) compared the survival of gamebird chicks in cereal fields with and without unsprayed headlands (the 6 m wide margin of the field). He found a substantially better survival of Pheasants Phasanius colchicus L. and Grey Partridge Perdix perdix L. in fields with unsprayed headland whereas the data for Red-legged Partridge Alectoris rufa L.were inconclusive. A correlation between abundance of chick food items and chick survival was found in Grey Partridge and Red-legged Partridge Alectoris rufa L. (Green 1984) and in Pheasant (Hill 1985). The significant impact of the unsprayed headlands on chick survival, despite their relatively small area, is explained by the preference of the investigated species for foraging in the field margin where more weeds and insects occur.
Experimental studies may provide insight into the relative importance of pesticide use to birds in cultivated fields and as such complement the bird surveys. From agricultural tests of pesticides it is well known that their efficacy in controlling pests varies between sites and years indicating that the magnitude of the pesticide impact will depend upon the weather and other environmental conditions. For purposes of bird conservation it is important to understand how the impact of pesticide use on the avifauna varies between years as well as to gain insight into the mechanisms behind this variation. A broader understanding of the ecology of different species and how their autecology determines their sensitivity to pesticide use, is valuable for bird conservation. This understanding allows us to predict the outcome of differing pesticide use scenarios, to assess the success of mitigating strategies, and to help the development of policies for the protection of the environment.
The present study is an attempt to quantify the impact of pesticide use on the arthropod food resource and the breeding success of Skylark Alauda arvensis L. Skylark nestlings are fed with various arthropods and the project builds on the hypothesis that prey availability is essential to breeding success. Food limitation may influence breeding birds by lowering quantity and quality of the offspring in a number of ways and the strategy of the parent birds in terms of how much effort to invest in a single brood may be determined by the life history of the species (Martin 1987).
The study was carried out in four fields subjected to different pesticide treatment allowing us to estimate the combined effect of herbicide and insecticide applications. As estimated by the use of analysis of variance, the average number of fledglings produced was substantially reduced when the fields were subjected to full pesticide treatment (Odderskær et al. 1997b). In the present paper the nestling food and the arthropod food supply in the fields subjected to different pesticide treatment are presented. The correlation within the breeding season between the breeding activity of Skylarks and the food resource are analysed with the aim of elucidating the mechanisms behind the effect of pesticide treatment.
The Skylark is characteristic of the arable land in large parts of Europe. It avoids tall vertical structures like trees etc. in the landscape, hence nesting and foraging takes place in the open fields. The species is therefore very exposed to agricultural operations in the fields.
In Denmark Skylark numbers declined from a breeding index of 100 in 1976 to 59 in 1986. Since then there has been an increase and the breeding index has fluctuated between 75 and 80 in the period from 1993 to 1996 (Jacobsen 1997). Petersen and Jacobsen (1997) did not find that pesticide use was a significant factor when modelling population development from 1976 to 1997. In UK lowland farmland, there was a decline of 51% between 1968 and 1995 (Siriwardena et al. 1998). A greater density of Skylarks has been reported in organically compared to intensively managed fields (Braae et al. 1988, Wilson et al. 1998).
The Skylark prefers low vegetation and forages in open patches within the vegetation (Odderskær et al. 1997b), hence the breeding density varies considerably between different crops. High densities are found in set-aside fields and spring cereals and low densities in dense and fast growing crops like winter wheat and winter oil seed rape (Wilson et al. 1998, Poulsen et al. 1998, Schläpfer 1988, Daunicht 1992).
METHODS AND STUDY AREA
The studies were conducted on Kal
r Estate, near Rrnde in Eastern Jutland, Denmark (56°8' N, 10°30' E) during 1992 - 1995. The design involved four fields, two sprayed and two unsprayed. Treatment switched every year during the four years.Fields were sown with the spring barley cultivar Canut. Due to a heavy infestation of mildew it was necessary to spray the ‘unsprayed’ fields with the fungicide 'Tilt Turbo' (see chemical names below) in 1993, 1994 and 1995 to avoid a significant loss of crop plant cover. Furthermore a seed dressing was used in both sprayed and unsprayed fields. Treatment is therefore referred to as Full Pesticide Treatment (FPT) and Reduced Pesticide Treatment (RPT) each area covering 50 ha.
Pesticides
The spring barley sown was treated with the seed dressing 'Fungazil' (emulsifiable concentrate containing 50 g imazalil l
)1, DLG) at a dosage of 5 mg a.i. per kg seed equivalent to 8.35 - 8.9 g imazilil ha-1.All sprayed fields were treated with the foliar fungicide 'Tilt Top' (emulsifiable concentrate containing 125 g propiconazole and 375 g fenpropimorph l -1, Ciba-Geigy) at a dosage of 0.5 l ha-1. Application dates were 18. May 1994 and 11. May 1995.
In 1993, 1994 and 1995 the 'unsprayed' fields were treated with 'Tilt Turbo' (emulsifiable concentrate containing 125 g propiconazole and 350 g tridemorph l -1, Ciba-Geigy) at a dosage of 1 l ha-1. Application dates were 14. June 1994 and 6. June 1995. Propiconazole has no insecticidal effects on most arthropods (Hassan et al. 1988; Sotherton and Moreby, 1988). Studies testing insecticidal effects of the active ingredient tridemorph showed no toxic effects on two insect species important as food items for game bird chicks (Sotherton 1989). In an experimental field study no negative effects on bird reproduction or on the abundance of potential bird prey items were found after spraying with 'Tilt Turbo' (Odderskær and Sell 1993).
The herbicide used in the sprayed fields was 'EK480' (emulsifiable concentrate containing 30 g dicamba, 500 g MCPA and 20 g clopyralid l-1, (Esbjerg Kemi) at a dosage of 2 l ha-1. Application dates were 18. May 1994 and 11. May 1995.
The insecticide 'Cyperb' (emulsifiable concentrate containing 100 g cypermethrin l-1, DLG) at a dosage of 0.25 l ha-1 was applied in the sprayed fields. Application dates were 6. June 1994 and 16. June 1995.
Nestling food
Nestling faecal pellets were collected at nest visits. When handled by the observer the nestlings usually defecated and all nestling faecal sacs produced during handling were collected. A maximum of six faeces per nest were analysed. Prior to analysis, the droppings were placed in a 20% sodium hydroxide solution for 20 minutes to remove uric acid and small particles which otherwise obscured the examination of the faeces. The faeces were then rinsed in distilled water in a glass filter crucible using a filter paper to collect the residue. After washing, the sample was transferred to a glass petri dish and arthropod fragments were identified under a binocular microscope (6 - 40 x magnification), by use of a reference collection of identified arthropods. As a minimum, arthropod fragments were identified to order, but most were identified to family, genus or species. The minimum number of individuals of one prey species in a pellet was counted. For all taxa one individual per faeces was assumed unless the number of mandibles, eyes, legs or wings recovered exceeded 2, 2, 6 (8 Araneae) or 4 (2 for Diptera), respectively. For adult butterflies the presence of many scales or hairs indicated that the birds had eaten a minimum of one individual.
Food abundance
Estimates of arthropod density were obtained by use of a Dietrick vacuum insect sampler (D-vac) (Dietrich 1961). As the sampling of ground beetles (Carabidae) by the D-vac is inefficient and biased (Sunderland et al. 1987), the Ground Search method (G-S) (Sunderland et al. 1987) was used as a supplement to estimate the abundance of carabids. As a measure of arthropod prey availability the two data sets were combined: G-S was used as ground beetle density estimates and D-Vac samples as a measure of density for all other taxa.
Ten D-vac samples were collected per sampling date on each field. Samples were taken at intervals of 7 to 14 days throughout the entire sampling period from May to July. All samples were collected between 14.00h and 21.00h, when the vegetation was dry. One sample comprised five sub-samples, each covering an area of 0.092 m2, and each lasting for 12-16 seconds. Sub-samples were taken 6 metres apart. Immediately after collection, the samples were transferred to an insulated bag and within a few hours frozen (-18°C) for storage. The samples were taken in different areas of the field in the vegetation bordering the tramlines. Each area included at least one skylark territory. Within each year samples were taken in the same areas throughout the breeding season. Prior to being sorted the samples were sieved. All samples were subsequently examined under a binocular microscope (6 - 40 x magnification).
Aphididae, Collembola, Thysanoptera, Acarina, Diplopoda, Opiliones and Chilopoda were not further identified. Diptera were divided into three groups: Nematocera, Tipulidae and Brachycera/Cyclorrhapha. All other orders were identified to family level and those orders known to be important as nestling food items were identified to genus or species level.
Fifteen G-S samples were taken within the same areas and time periods as the D-vac samples. The G-S samples were taken at 7 days interval in 1992 and with 14 day intervals in 1993, 1994, and 1995. One sample comprised three sub-samples each covering an area of approx. 0.19 m2. Sub-samples were taken by placing a circular metal ring with a diameter of 0.5 m at the edge of the tramline into the crop. The ring was pressed into the soil, and for 5 minutes the vegetation and the top soil were searched for ground beetles. All specimens were identified to species in the field except species of the genera Amara, Trechus and Bembidion which were identified to genus.
In the absence of information on the relative quality of food items, dry weight was used as a measure of food availability. Dry weight measures were obtained for most of the prey items. For prey species for which we did not have specific measures, extrapolation was made by comparing size and shape with species of known weight. Changes in weight of a species during season was not integrated.
Skylarks
Skylark territories were mapped on all fields several times a week from March to June following the method described by Enemar (1959) and Svensson (1970). In each mapping session, the observer covered the entire field on foot, noting also the mating status of territory owners. Between eight and 21 territories were identified in the individual fields during the four years (Odderskær et al. 1997b). Throughout the breeding season four observers were each responsible for a specific field. Count-efficiency was assumed to be independent of field and observer. In order to position birds within the fields, a grid of numbered flag-markers was set up in each field, dividing the fields into 18 m x 100 m sections.
Nests were located by observation of nest-building, systematic search in potential areas, flushing of incubating females from the nest, observations of adults feeding nestlings, radio-tracked females, or chance. When a nest was located, the exact position was transferred to field-maps, and its position in the field was marked with a thin plastic rod. Different coloured rods were used to indicate whether a nest was active at the nest building, egg or nestling stage. The majority of active nests (73%) were located during egg-laying or incubation. If located during egg-laying, the nest was checked every morning until the last egg was laid. From this day and 10 days onwards these nests were not visited. Nests located during incubation were checked every day until hatching. Most nestlings fledge at an age of 8 days. The age of nestlings in nests located during the nestling stage was estimated by aid of photographs of nestlings of known age.
Data analyses
Because temperature and arthropod density and species composition change considerably during the year, the season was divided in two periods: (1) from beginning of season until 14 days after insecticide application, (2) the rest of the season. For some analyses, period 1 was divided in 1a (until insecticide application) and 1b (the 14 days after insecticide application).
In analyses of diet composition, only orders accounting for more than 1% of the food items were included in a multivariate analysis of variance (% dry weight arcsin transformed) in sprayed and unsprayed fields) (PROC GLM/MANOVA; SAS Institute 1989). The contribution of each prey order to the difference between treatments was evaluated by canonical discriminant analysis (PROC CANDISC, SAS Institute 1989).
RESULTS
Nestling food
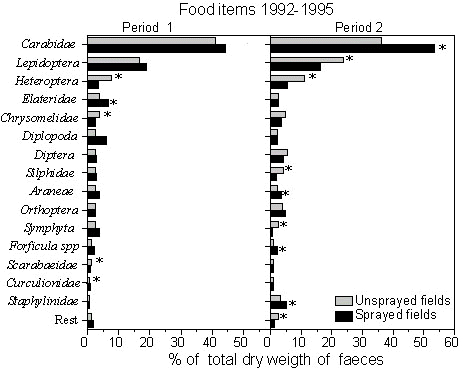
A total of 550 faeces were analysed; 249 from sprayed fields and 301 from unsprayed fields. Ground beetles were the predominant prey accounting for 42% of the estimated total dry weight of food items in the faeces. A maximum ground beetle content of 76% was found in period 2 in 1994 on sprayed fields. Butterfly imagines and larvae (Lepidoptera) (19%) and plant bugs (Heteroptera) (7%) were the only other orders contributing more than 5% to the total dry weight of food items. The composition of the faecal contents differed between nestling faeces from sprayed and unsprayed fields, Fig. 1.
The proportion of ground beetles did not differ between treatments in period 1, but in period 2 increased in faeces from sprayed fields. The same pattern was found for spiders (Aranea), earwigs Forficula spp., and rove beetles (Staphylinidae). Butterflies, Silphidae and Sawflies (Symphyta) occurred in equal proportion in faeces collected in sprayed and unsprayed fields in period 1, but in period 2 they were more frequent in unsprayed fields. Plant bugs and leaf beetles (Chrysomelidae) occurred more frequently in faeces from unsprayed fields both in period 1 and 2, although the difference in period 2 was not statistically significant (Fig. 1).
Food abundance
Arthropod species found in faeces were defined as food items and included in analyses of food abundance. Generally the food abundance, measured by use of D-vac and ground search, was very low in early May but increased during May and June.
The arthropod community was not only affected by pesticide treatment, it also differed between years and fields. In an analysis of variance of the D-Vac catches of food items (ln(dw)) with the main factors Treatment, Year and Field, all main factors came out significant as did Year*Treatment. The significant first order interaction invalidates the test of main factors. Only in the 14 days just after insecticide application (1b) were there no significant interactions and Treatment was significant as the only main factor (F1,11=37.55, P<0.004). From the ANOVA the food abundance in sprayed fields during this period was estimated at 36 % of unsprayed fields (95%; confidence interval: 21% to 51%). Due to the uncertainty of the density estimates it was rarely possible to detect differences in the abundance of single species between treatments on particular dates.
The occurrence of ground beetles in ground search samples was analysed in the same way as the D-vac data. No effect of Treatment was found (F1,11=0.91, P<0.40), but there was a significant effect of Year and Field (F2,11=35.18, P<0.01 and F2,11= 15.98, P<0.02 respectively). In 1992 the average carabid dry weight per 0.5 m2 per sampling was 10 mg, whereas in 1993 to 1995 it was 62 to 72 mg per 0.5m2 which indicate the difference between the years. The average dry weight per sampling in the fields during all 4 years varied between 27 and 79 mg per 0.5 m2 in the different fields.
Breeding activity and food abundance
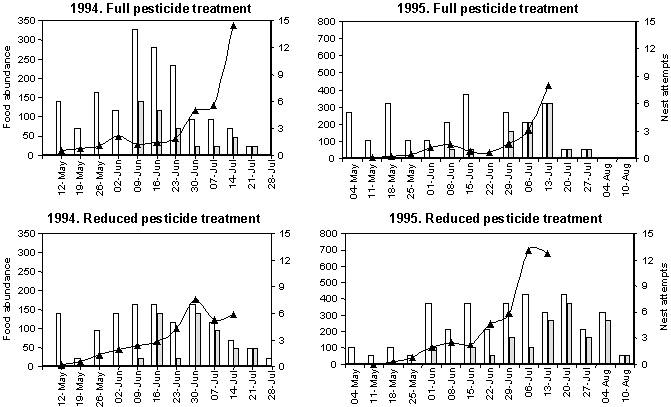
The critical size of the food supply depends on the food demand which, for a pair, increases when the eggs develop in the female and when they feed the nestlings. The number of nesting attempts initiated during a week was used as a measure of the energy invested in breeding and an indicator of increased food demand. In Fig. 2 data on food abundance and nest attempts are shown for 1994 and 1995. The figure illustrates some general results from all four years. A successful nest was defined as a nest where at least one fledgling was produced. Unsuccessful attempts were caused by predation, because nests were abandoned by the parents or all nestlings died presumably from starvation as no indication of illness was observed. If food abundance is a limiting factor for breeding success, it could be expected that breeding activity should be highest when the food supply is sufficient. The breeding activity, however, seemed unrelated to food abundance as the first breeding attempt is initiated at a time where there is a poor prey density and the success of the first breeding attempt is very low. The breeding season continues longer or the late season is more successful in RPT-fields. After some weeks the impact of the insecticide spraying in June (1994: 6. June; 1995: 16. June) on the weight of food items per unit area vanishes but the high food abundance in late season is not exploited in the FPT-fields. The food abundance in FPT fields during the late season in 1994 is primarily due to the hatching of large carabids Pterostichus melanarius Illig. from pupae in the soil. The larvae of P. melanarius live in the soil and are not exposed to significant insecticide concentrations.
In 1994 the number of young fledging was 44 and 59 in FPT-fields (50 ha) and RPT-fields (50 ha) respectively. In 1995 the corresponding numbers were 47 and 91.
DISCUSSION
The food supply in FPT-fields was depleted particularly in the weeks following insecticide treatment in June when the total weight of food items in D-vac samples on average was reduced by 64% compared to RPT-fields. Some important prey items, such as the ground beetles, were not significantly affected by insecticide applications probably due to confounding factors and the protected position of these organisms in the soil, below stones etc. during spraying. Ground beetle density differed markedly between years and fields and in the late season food abundance in some years could be greater in FPT-fields (Fig. 2). This was, however, not reflected in a greater Skylark reproduction during the same period. Based on analyses of faeces, the food of nestlings differed significantly between treatments. In FPT fields ground beetles were more predominant (Fig. 1). In RPT fields the food was more diverse: butterflies, plant bugs, and leaf beetles were more frequent in nestling faeces. In these herbivorous insect groups many species have host plants in the weed flora and they may benefit when herbicide application is omitted. Also, herbivores eating aerial parts of plants are very exposed to insecticides, directly or indirectly through their diet, and therefore often significantly affected by insecticide sprayings.
Dry weight was used as an indicator of the value of various prey species. It is a very coarse measure of food quality. Insects very rich in chitin, such as many beetles, have probably a lower quality per unit weight. Insects like caterpillars, which have a low chitin content are probably a better food source (Bell 1990; Kaspari and Joern 1993). Furthermore diversity in the food intake may be a quality in it self, making the nutritious value of a particular prey items dependent on what else is available. Insecticide application may not always deplete the food supply to a critical level for the birds, if food abundance still is above the food demand. Martin et al. (1998) studied the impact of insecticide application on the insect food resources and the performance of Chestnut-collared Longspurs Calcarius ornatus living in a grassland habitat. After spraying the diet changed significantly but nestling survival was unaffected. The alternative food items appeared equally good, some were even more available after spraying. In the study of the Chestnut-collared Longspurs a lower egg success in insecticide sprayed fields was observed as was the tendency in the present study (Odderskær et al. 1997b). This might be due to an effect of a change in the diet of females on the quality of the eggs produced.
The methods used to estimate food abundance in the present study are not ideal. The efficiency of the D-vac sampling is species specific and changes during the season as the vegetation develops. In late season when the crop is high, a number of insects found high in the crop are sampled although they are not available to the Skylarks, which exclusively forage on the ground. An accurate description of food density is difficult to make and precise estimates of food availability is hardly obtainable.
The relationship between food supply and breeding success in the Skylark within a single breeding season is hard to track for other reasons of much greater importance. The first breeding attempt occurs when the food resource is very poor and the risk of predation very high. The early breeding attempt may increased the fitness of the species on a larger temporal and spatial scale and should perhaps be measured in terms of life time reproductive success. The annual reproductive success, or the output from single breeding attempts, measured as number of fledglings, do not reflect the reproductive value of the young.
Late in the season while there is still plenty of food, breeding activity peters out in all fields because the prospects for offspring success are negligible. The breeding season of birds in FPT-fields was shorter than in RPT-fields, even when food supply was plentiful. This may be due to the poorer condition of the adult birds. At a given time the condition of the parent birds could be expected to be a function of food availability in the previous days or weeks and the time since the last breeding attempt (Martin 1987). Hence a time lag between food shortage and a lower breeding activity can occur and confound a simple correlation between food abundance and breeding activity.
A relationship between food supply and the number of broods has been observed in other bird species. The most significant impact of Bacillus thuringiensis treatment on Blue Warblers (Dendroica caerulescens) in a deciduous forest, was the lower number of nesting attempts (Rodenhouse and Holmes 1992). Simons and Martin (1990) found a higher prevalence of second clutches in Cactus wrens Campylorhynchus brunneicapillus provided with supplemental food.
In altricial birds the food demand of nestlings are satisfied via the parents. The food demand depends on age and size of the clutch, and the heat loss to the environment as a function of weather conditions. Weather also influences the time available for foraging of the parent birds. When it is cold and wet they have to spend more time on the nest. The significance of the amount of food available thus depends on the food demand and the time available for foraging. The mechanisms behind the indirect effects of pesticide on reproductive success in multi-brooded altricial birds may be complex and the impact very different from one year to another which emphasises the limitations of short term studies also in environmental research.
From the present four year study it can be concluded that herbicide and insecticide applications reduced the average breeding success markedly and the results suggests that a lower food supply is responsible for the observed effects, but the correlation between food supply and breeding activity within a season is not simple. It is possible that the effect of pesticide spraying on insect diversity particularly by reducing the number of herbivorous insects, may affect food quality as shown by studies on the Grey Partridge (Southwood and Cross 1969; Potts 1986). The indirect effects of pesticide use on the avifauna are related to the intended effects of the pesticide application, i.e. the reduction of weed and insects densities, although many of the affected insects are non-target species. The term non-target is not used for weed species although it might be appropriate for species of no economic importance. As the treatments with insecticides and herbicides in many cases can be viewed as generally non-specific towards arthropods and plants respectively, the magnitude of the indirect effects are correlated with the frequency of treatment. Thus the indirect effects cannot be met by contemporary legislative procedures and substitution of chemicals. Mitigating strategies have to involve a reduction in the frequency of treatments, more selective pest control programmes and the provision of untreated areas.
ACKNOWLEDGEMENT
Thanks to Dr. Pierre Mineau for improving an earlier draft of this paper.
REFERENCES
Bell, G.P. 1990. Birds and mammals on insect diet: a primer on diet composition analysis in relation to ecological energetics. Avian Biology 13: 416-422.
Braae, L., Nøhr, H. & Petersen, B.S. 1988. Fuglefaunaen på konventionelle og økologiske landbrug. Miljøprojekt nr. 102. København. Miljøstyrelsen.
Clausen, H. 1998. Ændringer i bekæmpelsesmidlernes egenskaber fra 1981 - 1985 frem til 1996. Roskilde. Danmarks Miljøundersøgelser. 63 s.
Daunicht, W.D. 1992. Einfluss der Bewirtschaftung am Beispil der Feldlerche. EcoSys 2: 73-74.
Dietrick, E.J. 1961. An improved backpack motorised fan for suction sampling of insect populations. Journal of Economic Entomology 54(2): 394-395.
Enemar, A. 1959. On the determination of the size and composition of a passerine bird population during the breeding season. Vår Fågelvald 2: 1-114.
Green, R.E. 1984. The feeding ecology and survival of partridge chicks (Alectoris rufa and Perdix perdix) on arable farmland in East Anglia. Journal of Applied Ecology 21: 817-830.
Hald, A., Reddersen, J. 1990. Fugleføde i kornmarker - insekter og vilde planter. København. Miljøprojekt nr. 125. Miljøstyrelsen.
Hassan, S.A., Bigler, F., Bogenschutz, H., Boiler, E., Brun, J., Chiverton, P., Edwards, F., Mansour, F., Naton, E., Samsøe-Petersen, L., Stäubli, GA., Sterk, G., Tavares, K., Tuset, J.J., Vigianni, G. & Vivas, A.G. 1988. Results of the fourth joint pesticide testing programme carried out by the IOBC/WPRS-working group 'Pesticides and beneficial organisms. Journal of Applied Entomology. 105: 321-329.
Hill, D.A. 1985. The feeding ecology and survival of Pheasant chicks on arable farmland. Journal of Applied Ecology 22: 645-654.
Jacobsen, E.M. 1997. Monitering af agerlandets fugle 1995 og 1996. København. Miljø- og Energiministeriet, Miljøstyrelsen.
Kaspari, M. & Joern, A. 1993. Prey Choice by Three Insectivorous Grassland Birds - Reevaluating Opportunism. Oikos 68(3): 414-430.
Martin, T.E. 1987 Food as a limit on breeding birds: life-history perspective. Annual Review of Ecology and Systematics 18: 453-487.
Martin, P.A., Johnson, D.L., Forsyth, D.J. & Hill, B.D. 1998. Indirect effects of the pyrethroid insecticide deltamethrin on reproductive success of chestnut-collared longspurs. Ecotoxicology 7: 89-97.
Moreby, S.J., Aebischer, N.J., Southway, S.E. & Sotherton, N.W. 1994. A comparison of the flora and arthropod fauna of organically and conventionally grown winter wheat in southern England. Annals of applied Biology 125: 13-27.
Odderskær, P., Prang, A., Poulsen, J.G., Andersen, P.N. & Elmegaard, N. 1997a. Skylark (Alauda arvensis) utilisation of micro-habitats in spring barley fields. Agriculture, Ecosystems & Environment 62: 21-29.
Odderskær, P., Prang, A., Elmegaard, N. & Andersen, P.N. 1997b. Skylark Reproduction in Pesticide Treated and Untreated Fields. Pesticide Research 32. Copenhagen. Danish Environmental Protection Agency.
Odderskær, P. & Sell, H. 1993. Survival of great tit (Parus major) nestlings in hedgerows exposed to a fungicide and an insecticide: a field experiment. Agriculture, Ecosystems & Environment 45: 181-193.
Petersen, B.S., Falk, K. & Bjerre, K.D. 1995. Yellowhammer Studies on Organic and Conventional Farms. Pesticide Research 15. Copenhagen. Danish Environmental Protection Agency.
Petersen, B.S. 1996. The distribution of birds in Danish farmland. Pesticide Research 17. Copenhagen. Danish Environmental Protection Agency.
Petersen, B.S. & Jacobsen E.M. 1997. Population Trends in Danish Farmland Birds. Pesticide Research 34. Copenhagen. Danish Environmental Protection Agency.
Potts, G.R. 1986. The Partridge: pesticides, predation and conservation. London, Collins.
Poulsen, J.G., Sotherton, N.W. & Aebischer, N.J. 1998. Comparative nestling and feeding ecology of skylarks Alauda arvensis on arable farmland in southern England with special reference to set-aside. Journal of Applied Ecology 35: 131-47.
Rands, M.R.W. 1986. The survival of gamebird (Galliformes) chicks in relation to pesticide use on cereals. Ibis 128: 57-64.
Reddersen, J. 1997. The arthropod Fauna of Organic Versus Conventional Cereal Fields in Denmark. Biological Agriculture & Horticulture 15: 201-211.
Rodenhouse, N.L. & Holmes, R.T. 1992. Results of experimental and natural food reductions for black-throated blue warblers. Ecology 73(1): 357-372.
SAS Institute. 1989. SAS/STAT User's Guide, Version 6 fourth edition. Cary NC, SAS Institute Inc.
Schläpfer, A. 1988. Populations Ökologie der Feldlerche Alauda arvensis in der intensiv genutzten Agrarlandschaft. Der Ornithologishe Beobachter 85: 309-371.
Simons, L.S. & Martin, T.E. 1990. Food limitation of avian reproduction: An experiment with the Cactus Wren. Ecology 7(3): 869-876.
Siriwardene, G.M., Baillie, S.R., Buckland, S.T., Fewster, R.M., Marchant J.H. & Wilson, J.D. 1998. Trends in abundance of farmland birds: quantitative comparison of smoothed Common Birds Census indices. Journal of Applied Ecology 35: 24-43.
Sotherton, N.W. 1989. Farming Practices to Reduce the Exposure of Non-target Invertebrates to Pesticides. Pesticides and non-target invertebrates, ed. Jepson, P.C. Intercept, Dorset: 195-212.
Sotherton, N.W. & Moreby, S.J. 1988. The effect of foliar fungicides on beneficial arthropods in wheat fields. Entomophaga 33(1): 87-99.
Southwood, T.R.E. & Cross, D.J. 1969. The ecology of the partridge. III. Breeding success and the abundance of insects in natural habitats. Journal of Animal Ecology 38: 497-509.
Sunderland, K.D., Hawkes, Q., Stevenson, J.H., McBride, W., Smart, L.E., Sopp, P.I., Powell, W., Chambers, R.J. & Carter, O.C.R. 1987. Accurate estimation of invertebrate density in cereals. Bulletin SROP 10: 71-81.
Svensson, S. 1970. Recommendation for an international standard for a mapping method in bird sensus work. Acta Ornithologica 14: 322-338.
Wilson, J.D., Evans, J., Browne, S.J. & King, J.R. 1998. Territory distribution and breeding success of skylarks Alauda arvensis on organic and intensive farmland in southern England. Journal of Applied Ecology 34: 1462-1478.
Fig. 1. Composition of arthropod food items (% of total dw) in nestling faeces from fields with full (sprayed) and reduced pesticide treatment (unsprayed), 1992 - 1995. Period 1: Until 14 days after insecticide spraying. Period 2: from 14 days after insecticide spraying and the rest of the sampling season. Asterisks indicate significant difference (a=0.05) between sprayed and unsprayed fields tested by use of MANOVA and Canonical discriminant analysis.
Fig. 2. Food abundance (mg dw 0.5 m-2 ) (line), total number of breeding attempts (filled bars), and unsuccessful nest attempts (no fledglings)(empty bars) during the breeding season 1994 and 1995 in fields with full (FPT) and reduced pesticide treatment (RPT)