S20.4: Whose young win? Parental quality and recruitment in seabirds
Peter H. Becker
Institut für Vogelforschung 'Vogelwarte Helgoland', An der Vogelwarte 21, D-26386 Wilhelmshaven, Germany, e-mail p.becker@ifv-terramare.fh-wilhelmshaven.de
Becker, Peter H. 1999. Whose young win? Parental quality and recruitment in seabirds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1190-1208. Johannesburg: BirdLife South Africa.Measuring fitness is especially difficult in long-lived seabirds. Available information shows that only a small percentage of breeders will have offspring recruiting to the breeding population. Most losses to breeding age occur during the long post-fledging stage. The paper addresses the question which characteristics qualify a fledgling to become a recruit by comparing natal characteristics. Besides reviewing the literature some preliminary data from a new methodological approach at a colony of Common Terns Sterna hirundo marked with transponders are presented. In several species, date of fledging, number of fledglings per brood and their body mass were found to affect survival to recruitment, whereas hatching position and parental age mostly were found not to be important after fledging. Another research possibility is to investigate breeders longitudinally to identify traits which predict the number of recruits. Longevity, high parental quality and body condition, and also a high number of fledglings per year are found to be important, often concomitant traits enhancing fitness. Thus, a state-dependent approach to life-histories of seabirds is supported. Individual quality is one key for understanding fitness in long-lived species like seabirds.
INTRODUCTION
Fitness, or the number of reproductive offspring of an individual organism, is an important parameter in population ecology. Animals produce much more offspring than ever will reproduce themselves. Most young die by random events, environmental constraints and competition for resources, and are excluded from reproduction. Or, when becoming a breeder, they may be of low quality, not able to produce surviving young. In this way natural selection is preventing the majority of individuals from contributing young to the next generation.
In long-lived birds only a small proportion of the population provides recruits for the next generation (reviews in Newton 1989), but only a few seabird studies were able to record fitness directly: In Red-billed Gulls Larus novaehollandiae, only 17% of males and 24% of females which bred produced young which survived to breed (Mills 1989). In Short-tailed Shearwaters Puffinus tenuirostris about one third of the breeders is responsible for most of the next generation (Wooller et al. 1989); 62% of fledged Kittiwakes Rissa tridactyla never had own fledglings; 38% of the overall variance in lifetime reproductive success could be attributed to pre-breeding mortality, a further 33% arose from variation in breeding life-span (Thomas & Coulson 1988). Those Kittiwakes with higher survival were characterized by higher individual quality and fledged more young per year and during their lifetime.
These data show that, besides strong variation among breeders in individual quality, it is mainly the survival of offspring to first breeding that determines the number of reproducing young of a seabird: Only 8% of Red-billed Gulls (Mills 1989) and 9% of Common Gulls (Larus canus, Rattiste & Lilleleht 1986) survive from egg to recruitment stage. Only 15-19% of the fledged chicks of Short-tailed Shearwaters which had bred once recruited to their natal colony (Wooller et al. 1990), and about 25% in the Manx Shearwater (Puffinus puffinus, Brooks 1990). In Western Gulls (Larus occidentalis) the figure is 22-24% (Spear & Nur 1995), in Roseate Terns Sterna dougallii 16% (Spendelow 1991), and in Little Blue Penguins (Eudyptula minor, Dann & Cullen 1990) 31% of the fledged young will reach breeding status.
Whereas factors influencing survival during the natal stage are well known in seabirds, those during the post-fledging stage up to recruitment are poorly investigated owing to methodological problems, despite the higher loss rates of offspring during this crucial phase, which is an important objective for operation of selective ecological and also microevolutionary factors (Cooke et al. 1991). Whether a bird survives to breeding age may depend on unpredictable environmental events, but also on direct or indirect effects of parental breeding performance and effort, including post-fledging parental care in some seabird species (Burger 1980), and on genetic factors (Cooke et al. 1991).
Our knowledge of the traits that characterise successful recruits and of the parents which produce them is still poor. Seabirds are qualified to be studied in this respect from several reasons:
- The high degree of philopatry and colonial breeding in great numbers facilitates identifying recruiting natal young, which is a prerequisite to relate parents’ reproductive performance with offspring survival. On the other hand, longevity and delayed first breeding cause methodological problems and require logistically expensive long-term studies (e.g. Bradley et al. 1990).
- In long-lived birds like seabirds, year effects which may obscure selection on characters in short-lived species (Price & Boag 1987), are of lesser importance. As consistency over years can be evaluated, inter-individual variation in fitness components like reproductive success and recruitment is easier to identify. Whether adult quality, which is related to survival (e.g. Coulson & Thomas 1988), may be age- or state-dependent (Forslund & Pärt 1995; McNamara & Houston 1996; Wendeln & Becker in press) is recently in discussion and may get further input from seabird studies in future;
- Many seabird species perform post-fledging care (Burger 1980), which may affect post-fledging survival besides inherited characters or influences from parental performance.
In this paper I address the following questions:
(1) What is known on the factors determining survival from fledging- to recruitment-age in seabirds? From the fledgling perspective, I review possible effects of natal characteristics like parental age, date of fledging, body mass at fledging, fledglings per brood, hatching order, and post-fledging care. The starting point is to compare survival probabilities from fledging to recruitment, or to compare traits between natal young re-encountered prospecting or recruiting and those not re-encountered.
Spear and Nur (1994) showed in their analyses of pre-breeding mortality of Western Gulls that fledging success is a poor indicator of the recruitment probability of a hatchling because of selection occurring post-fledging and influenced by factors mentioned above. Consequently also fitness-calculations may be problematic if based on lifetime reproductive success alone. Therefore direct relationships between individual parents, their quality and survival of their young to recruitment are needed:
(2) What traits of seabird parents predict the number of recruiting offspring? In longitudinal studies individual parents are followed over a life stage or over the total life-span to assign a fitness value to each individual (Price & Boag 1987). Fitness may be enhanced by genetic components and/or by phenotypic characters like e.g. size, or by a breeder’s reproductive effort and parental care. In some parameters, its consequences may affect the survival of fledglings in a direct way (see 1). Another area of interest is the intergenerational transmission of reproductive traits.
Besides reviewing the literature, I report first preliminary data gathered by a new methodological approach on Common Terns Sterna hirundo. Since 1992 some adults and all fledged young have been marked by passive transponders (Becker & Wendeln 1997). The methods used allow us to record not only the marked breeders but also the numerous prospecting pre-breeders, their body condition, fate and reproduction at the colony site for lifetime.
METHODS
I use definitions according to Danchin et al. (1991) and Bradley & Wooller (1991).
Prospector is restricted to immature natal prospectors, recruit to natal recruits breeding for the first time.
To study survival probabilities from fledging to return to breed, most authors compare factors between natal young re-encountered prospecting or recruiting and those not re-encountered. Birds not recovered as natal recruits include individuals which did not survive, were not detected or not philopatric. Unfortunately recruitment to other colonies in most studies remains unexplorable. Because of the high immature mortality, however, the majority of the non-recruited sample usually can be assumed not to have survived to recruitment and verifies this approach.
The Common Tern population study at Wilhelmshaven
The colony site is situated in the Banter See, a brackish lake in the harbour area of Wilhelmshaven, German North Sea coast. During the study period 1992-1997, the colony increased from 90 to 200 pairs breeding on six artificial islands of concrete ("closed" colony; details in Becker 1996; Becker & Wendeln 1997). From 1992-1995, 101 adults were caught and equipped with subcutaneously injected passive transponders sending an individual alphanumeric code when activated by special antenna (Becker & Wendeln 1997). In this way, a bird can be identified during the whole life without retrapping. The age of many adults was known because all chicks of this colony have been ringed since 1980. All fledged chicks of each year-class were fitted with a transponder. The transponders are read by antennas placed successively around all nests or on resting sites equipped with electrical balances to record the body mass of many of the marked terns. For the breeders weighed a daily average mass value was calculated, which was the basis for mean mass-values over the incubation and chick rearing period (for details see Wendeln & Becker 1996, Wendeln & Becker in press). Average body mass of a prospector was calculated if it was weighed at least for three times. Also arrival dates of breeders and pre-breeders are known.
Every two to three days, checks of reproductive biology were performed. Each nest and egg with respect to laying sequence was marked. All newly hatched chicks were ringed, and in most broods hatching sequence was known. Chick growth rates (3 - 13 d), peak and fledging mass were recorded weighing the ringed chicks at five of the six islands. Data used for this paper are laying date (5 d periods, also corrected for year effects), hatching date, hatching order, growth rate and peak mass of chicks.
Breeding success of this colony was very high during 1992-1995 (1.6 chicks per pair and year). Adult mortality was on average 11%, subadult survival to age of two years 33% (varying from 22 - 46% among years, Wendeln & Becker 1998). From the cohorts 1992-1994, 128 (31%) of 409 fledglings had prospected and 60 (15%) had recruited the colony until 1997. In 1997 e.g., 132 prospectors and 39 recruits were present. Until 1997, a total of 209 natal prospectors were recorded from which 61 birds had recruited to the natal colony.
Most natal immatures prospect the colony two years old (76%, Wendeln & Becker 1998). To study prospecting including 1998, I used cohorts of young and parent data from 1992 - 1995. Data include parents between 3 - 15y. Birds with complete data sets for three breeding seasons (14 males, 17 females) fledged 2 - 9 chicks, of which 0 - 4 became prospectors. First recruiting of most immatures starts at age three; correspondingly, the analysis of recruitment until 1997 is based on cohorts and adult data from 1992 – 1994. Some more prospectors from these cohorts are expected to recruit after 1997, and the data base still is relative small, that multifactorial analyses are not yet possible. Therefore the results presented on Common Terns should be regarded as preliminary. Average values are given as means ± S.D.
RESULTS
Post-fledging survival and condition of prospectors or recruits in relation to parental age and natal characteristics
Influence of parental age and experience
In Short-tailed Shearwaters a significantly higher proportion of fledglings from first time breeders survived to recruitment than of fledglings from experienced breeders (Wooller et al. 1990). Thereafter the proportion of young which returned to breed increased only slightly with an increase in breeding experience. The proportion of young Kittiwakes fledged and returned to breed was independent of the previous parental breeding experience (Wooller & Coulson 1977). Also Nisbet (1996) found no parental age differences between Common Terns encountered and those not re-encountered breeding.
Influence of fledging date
Nisbet (1996) found no significant differences in hatching date of Common Terns that were encountered breeding and those not re-encountered. I also did not find a relationship between date of hatching and prospector rate in the total data set of 1992-1995 (rs = -0.01, n.s., n = 447). Testing the four years separately, only in 1992 the prospector rate decreased with delayed hatching (rs = -0.35, p < 0.01, n = 63). Also the cohort-dependent differences in immature survival (Wendeln & Becker 1998) were not related to the annual variation in the laying date.
Also in Guillemots Uria aalge only in some of the years studied early hatching had a positive effect on immature survival (Harris et al. 1992, two of six years; Hedgren 1981: no effect on first year-survival of five year classes). Neither Harris & Rothery (1985) found any effects of fledging date on the post-fledging survival of young Puffins Fratercula arctica, nor Lloyd (1979) in Razorbills Alca torda.
In Western Gulls Larus occidentalis Spear & Nur (1994) found decreasing survival probability with increasing hatching date in the juvenile stage; a comparable but weak trend was detected in the subadult stage. Also the recruitment probability was influenced in the same way by the hatching date. In the Herring Gull Larus argentatus higher post-fledging survival was found in early hatched young (Nisbet & Drury 1972, Parsons et al. 1976: in two of three cohorts), also in Black-headed Gulls Larus ridibundus (Viksne & Janaus 1993).
Late fledged Manx Shearwater Puffinus puffinus chicks suffered higher mortality, but post-fledging survival was also affected by the weight of the chicks which are lighter when hatching late in the season (see above; Perrins 1966; Perrins et al. 1973). Thus, timing of breeding ultimately affects the survival of the young. Also in African Gannets Sula capensis (Järvis 1974) early date of fledging had a positive influence on survival.
Hatching order and fledging success
In seabirds laying more than one egg and rearing more than one chick, brood size and hatching order are known to influence chick survival, growth and body mass in various ways (e.g. Spear & Nur 1994; Nisbet et al. 1995). Some studies address the question, whether these parameters also affect the fate of the young after fledging.
In Herring Gulls Nisbet & Drury (1972) and Parsons et al. (1976) showed that young from broods of three fledglings had a lowered survival probability (in Parsons et al. 1976 only in one of 3 years, however). Owing to the results of Nisbet & Drury (1972), Herring Gulls had no discernible tendency for the third chick to suffer greater postfledging mortality. In contrast, Black-headed Gulls surviving the post-fledging period came from broods with more fledglings (2.3) than the average (1.2; Viksne & Janaus 1993). Kittiwakes (a- and b-chicks) from broods of three survived better than from broods of two fledglings, and a-chicks in broods of two survived twice as well as the b-chicks (Coulson & Porter 1985). In the Western Gull, brood size covaried positively with juvenile survival, but not with survival in the subadult stage and with recruitment probability (Spear & Nur 1994). In this species hatching order only had weak negative effects on juvenile survival. No effects of hatching order were detected in the Black-headed Gull (Viksne & Janaus 1993).
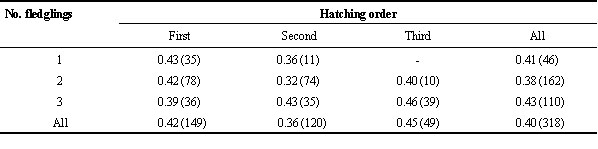
In the Common Tern neither the number of fledged siblings nor their hatching order had significant effects on their survival to prospecting (Table 1; see also Nisbet 1996). We found a weak tendency that arrival date of prospectors in the first season they returned to the birthplace may be influenced by hatching position: First chick 177 ± 18 (day of the year), n = 65; second chick: 181 ± 15, n=44; third chick: 185 ± 16, n=23; Kruskal-Wallis test, c 2 = 4.23, P = 0.12).
Chick mass and post-fledging survival
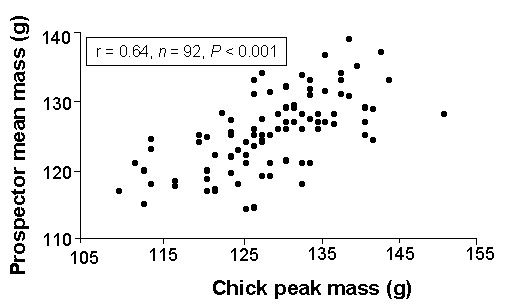
In the Common Tern growth rate (r = 0.27, n = 91, p < 0.01), and even stronger peak chick mass (r = 0.64, P < 0.001, n = 92, Fig. 1) are positively correlated with the chicks’ later mass during the period of prospecting at the natal colony site. Does the individuals’ mass influence its prospecting or recruiting rate? The data do indicate only small effects: Common Tern chicks which fledged and returned as prospectors had only slightly higher peak body mass (129 ± 8 g, n = 132) than individuals which did not return (127 ± 8 g, n = 212; t-test, t = -1.92, p = 0.06). No effects of fledging mass on immature survival were found in Guillemots (Hedgren 1981, Harris et al. 1992), Puffins (Harris & Rothery 1985) or Razorbills (Lloyd 1979).
In the Manx Shearwater Puffinus puffinus immature survival increased with peak chick mass (Perrins et al. 1973), also in African Gannets Sula capensis (Jarvis 1974) young dying soon after fledging were lighter than the average. The proportion of Kittiwake chicks which survived and returned as recruits increased significantly in relation to their growth rates as chicks (Coulson & Porter 1985).
Effects of condition on probability and age of recruitment
In Kittiwakes of both sexes recruits were heavier than prospectors (Porter & Coulson 1987). Only small effects of body condition on return rates were found in Common Terns (see above), and also the probability to recruit seems not to be influenced by body mass: Neither chick peak mass nor mass at prospecting differed dependent on recruitment in the subsequent year (chick peak mass: Prospectors not recruiting in the subsequent year 127 ± 6 g, n = 33, prospectors recruiting the next year 129 ± 10 g, n = 38; t = -0.89, P = 0.38; prospector mass: 125 ± 6 g, n = 35; 125 ± 5 g, n = 36; t= -0.33, P = 0.74).
Interesting relationships, however, were found between the age of first breeding, body mass and date of prospecting in the year when a chick first returned (Table 2). Age of recruitment and chick peak mass were negatively correlated (rs = -0.42, p < 0.01, n = 39). Young recruiting at age three had higher peak mass when chicks than those which recruited when 4 years or older. The latter tended also to have lower mass when prospecting (Table 2). Young which commenced to breed at age two had the highest peak mass. Moreover, age of recruitment was also related to the date when a prospector arrived at the colony site for the first time: Birds recruited at older ages had arrived as immatures on a later date (Table 2). Similarly recruiting Guillemots had arrived in the previous season earlier than birds not recruiting in the subsequent season (Halley et al. 1995).
Parent quality and characteristics of parents producing prospectors or recruits
Relationships between longevity, number of young produced and recruitment
Two seabird species have been studied in this respect. Red-billed Gulls were investigated over their total life-span by Mills (1989). Those birds which produced a large number of fledglings tended to have more progeny recruited into the population than less productive individuals, for both females and males, even if weaker correlated in the latter. Male and female longevity accounted for about 5% of the variance in the production of natal recruits. In Short-tailed Shearwaters Wooller et al. (1989) found a clear positive relationship between the number of young fledged by a bird during its lifetime and the number of reproducing offspring which it produced. Breeding life-span was positively correlated with the mean number of reproducing offspring.
In the Common Tern we can as yet consider only a section of one to three breeding periods of the individual life-span. Nevertheless, similar positive correlations between the number of fledglings, prospectors and recruits of the individual male or female birds were found (Table 3) as in Red-billed Gulls and Short-tailed Shearwaters.
Post-fledging survival rate was not related to the quality of Red-billed Gull parents of either sex as measured by the total number of young they fledged (Mills 1989). In Common Terns, too, the prospector rate itself did not depend on the number of fledged chicks (mean number of prospectors/mean number of fledglings per individual; males: rs = 0.08, n = 20, P = 0.75; females: rs = 0.18, n = 27, P = 0.38; only individuals recorded for at least three breeding seasons). These findings show that the chance to survive the post-fledging period seems not to be influenced by the number of fledglings a parent has to care for. Also the chick perspective (see above) indicates a similar amount of care spent per fledgling, or even an increasing care with fledgling number in gulls (Table 5). To achieve this, parents attending more than one fledgling have to increase their effort per brood.
Influence of body condition and laying date of parents?
Only two studies have adressed the question whether the body mass of seabird parents affects the number of recruits. Mills (1989) showed that Red-billed Gull which were heavier indeed had more fledglings, but not more young which survived to breeding age than lighter parents. In the Common Tern, mass which is largely independent from size (Wendeln et al. 1997) has a strong influence on chick growth and reproductive success (Wendeln & Becker in press). That female body mass is also positively related to the number of prospecting offspring shows the still small data base (Fig. 2, Table 4).
The individual laying date influences not only lifetime productivity (e.g. Red-billed Gull, Mills 1989), but also the lifetime number of recruiting offspring. In female Red-billed Gulls it was negatively correlated with laying date (not significant in males, Mills 1989). In Common Terns, first data show, that mean individual onset of laying and of egg-volume which depend on body mass (Wendeln 1997a) but not on size (Gonzaléz-Solís, Becker, Jover & Ruiz, unpubl. ms) do not correlate significantly with the number of natal prospectors of an individual (Table 4).
The consequences having well fed chicks
In the Common Tern I found a positive relationship between the mean peak mass of fledged chicks in a brood and the mean mass of the later prospectors from these broods (r = 0.62, p < 0.001, n = 39 broods). Male Common Terns rearing fledglings with on average higher peak mass had also more recruits (rs = 0.57, P < 0.05, n = 18; females: rs = 0.23, P = 0.26, n = 25; birds studied for at least two years), but not more prospectors (rs = 0.03, P = 0.92; females: rs = 0.15, P = 0.46; see also Table 4). These results correspond to that for individual Common Tern chicks (see above, Fig. 1) and show, that feeding performance of parents may affect the probability of survival and recruitment age of their offspring.
Importance of parental age and experience
The first data of the Common Tern in Wilhemshaven do not indicate age effects on the number of prospectors or on survival rate to prospecting (Table 4). However, age effects may be most pronounced in the very early stages of the reproductive life, of which yet only few data are available.
DISCUSSION
The information on factors affecting post-fledging survival in seabirds presented above is summarised in Table 5. Similarities, but also differences between species become obvious.
Direct effects on young
Deferred fledging date has negative consequences on post-fledging survival in the majority of species, even if in three species only in some years. Harris et al. (1992) suggest that post-fledging survival in Guillemots may be negatively affected by date only in favourable breeding seasons. This was not likely in the Common Tern at Wilhelmshaven: An effect of fledging date was found only in 1993, when an average breeding success was reached. The survival advantage of hatching early in the season may reflect direct effects (Spear and Nur 1994). Chicks fledging early may establish dominance over other chicks, or may profit from the longer time period to get experience in foraging before migration and before confronted with the strong environmental conditions in winter. But also indirect effects of having parents of better quality for which early laying is an indicator (e.g. Red-billed Gull, Mills 1989; Common Tern, Wendeln 1997a), may in part cause the advantages of fledging early (but see below).
Both direct effects and indirect parental influences may also cause the advantages of fledging together with siblings in species rearing more than one chick. Interactions between siblings may have positive effects on their competitive abilities (Spear and Nur 1994). The absence of effects in Herring Gulls and Common Terns show, that possible disadvantages to have siblings don’t exist or are at least outweighed by the advantages. As parents which are able to rear bigger broods again are of good quality (Thomas & Coulson 1988; Wendeln & Becker in press), the positive effects of the number of fledglings may reflect parental quality, too.
Fledging mass had positive influences on post-fledging survival except in the alcid species, which continue to grow after fledging (Gaston 1985). Consequently, Harris & Rothery (1985) warn of using mass at fledging as an indicator of parents’ breeding success in alcids. Body mass at independence, however, might influence a chicks survival prospects (Harris et al. 1992). In other species, heavier chicks will profit from the higher body reserves in the post-fledging period, when they leave the colony and parental influence ceased as in Manx Shearwaters (Perrins et al. 1973). Fat reserves may give young a greater survival chance during migration. Cooke et al. (1991) found a higher fledgling body condition index in recruiting than in non-recruiting Lesser Snow Geese Anser caerulescens caerulescens, showing directional selection between fledging and recruitment favouring larger birds. In the Common Tern, I found only a weak positive influence of fledgling mass on survival to prospecting.
Porter (1988) suggested, that differences in individual quality of Kittiwakes cause the variation in the age of first breeding and the ability to recruit the colony. Heavier weight may assist earlier arrival and increased colony attendance, first when prospecting and later when recruiting. Also Halley et al. (1995) hypothesised that earlier recruitment may be a secondary reflection of the underlying quality of individual Guillemots. Supporting these assumptions, fledgling peak mass was related to the age of recruitment and to the date of the first return of immature Common Terns to their natal colony (Table 2). The data also indicate direct relationships between these mentioned parameters and prospector mass, which can be studied by our methods. These now allow detailed monitoring of the physiological condition of prospectors which can be related to their prospecting pattern and recruitment parameters (Boulinier et al. 1996). Thus, dependence of the age of first reproduction on condition which is described in mammals (McNamara & Houston 1996) is indicated by our preliminary data in the Common Tern.
Except for the Kittiwake, hatching order had no influence on post-fledging survival in the studied seabird species. In the natal stage third chick disadvantage is well known in Laridae and other seabirds (e.g. Spear & Nur 1994). When fledged, however, third chicks have a similar chance of surviving to prospecting stage than first or second chicks. The selection processes during the natal period may let the c-chicks of best quality survive, and that is possible only in seasons of high food availability (Becker unpubl.); in Common Tern broods of three fledglings, c-chicks even show a tendency (n.s.) towards better survival to prospecting than a-chicks (Table 1).
Breeding experience of the parents was not shown to be an important factor for post-fledging survival (Wooller & Coulson 1977; Nisbet 1996) as proved for reproductive parameters (e.g. Clutton-Brock 1988). One reason may be the dominating importance of parental quality (e.g. Thomas & Coulson 1988).
Parental quality
Some of the factors discussed above which affect post-fledging survival may be ultimately affected by the parents’ quality. Direct or indirect parental effects, however, are difficult to disentangle. Longitudinal studies (summarised in Table 5) allow us to assign a fitness value to each individual parent (Price & Boag 1987). In this way phenotypic traits of individuals can be tested as to whether they are correlated with fitness components like the number of surviving offspring.
Spear & Nur (1994) showed in their analyses of pre-breeding mortality of Western Gulls that fledging success is a poor indicator of the recruitment probability of a hatchling, and consequently of an individuals’ fitness, because of selection occuring post-fledging and influenced by factors discussed above. Therefore a measure closer to fitness would be desirable, like survival to age one year (Western Gull, Spear & Nur 1994), or survival to prospecting like in the Common Tern. Such paramaters are difficult to record, however. The finding that immigrants and natal recruits do not differ in size, breeding performance or demography (Kittiwakes, Wooller & Coulson 1977; Short-tailed Shearwaters, Bradley et al. 1989) validates the approach to link parental traits with that of their natal recruiting young, which may be not biased because of their philopatry. As many breeders never have fledged young (e.g. Kittiwake: 11% of males, 8% of females, Coulson & Porter 1985; Red-billed Gull: 36% of males, 39% of females, Mills 1989) the best approach would be to take the number of recruits that successfully reproduce as measure of fitness, taking the reproductive value of offspring into account (McNamara & Houston 1996).
Longevity and number of offspring produced
Owing to delayed age of recruitment only a few studies have enabled us to relate parental reproductive traits and quality in seabirds with recruitment of their young. These studies correspondingly show the importance of longevity and of the number of chicks fledged for recruitment. Also in Little Blue Penguins (Dann & Cullen 1990), Fulmars Fulmarus glacialis (Ollason & Dunnet 1978) and Kittiwakes (Coulson & Thomas 1985), breeding lifespan accounts for much of the variability in lifetime reproductive success. Wooller et al. (1990) and Weimerskirch (1992) describe discrete age-stage dependent trade-offs between survival and fecundity to increase the reproductive value; by that long-lived seabirds can maximise their lifetime offspring production.
Post-fledging care and food provisioning
Post-fledging care is extended to more than one month after fledging in cormorants, boobies, frigatebirds, gulls and terns, especially in species raising more chicks, to increase the survival prospects of the young (Burger 1980). Care after fledging may support the strategy to produce more than one fledgling in order to enhance their recruitment prospects. Also the positive direct effects of siblings on fledglings (see above) point to selection favouring more-fledgling broods.
Not only the number of fledglings a Common Tern parent raises per season, also their condition seems to be important for recruitment. Mainly male Common Tern parents share the duties of food provisioning of young (Wiggins & Morris 1987; Wendeln 1997b), and initial data indicate that males rearing broods with well fed chicks had more recruiting offspring. Chicks characterised by higher peak mass recruit at a younger age than birds that are lighter during the rearing period. In the Kittiwake, Porter (1988) reported similar relationships.
Age, experience
In many seabird species reproductive performance and breeding success increase during the first breeding years (Kittiwake and Common Tern 3-4 years, Coulson & Thomas 1985; Nisbet et al. 1984; Fulmar 9 years, Ollason & Dunnet 1988; Clutton-Brock 1988). However, often quality effects explain more of the variation than age effects, as e.g. in Kittiwakes (Thomas & Coulson 1988) or in egg size of Wandering Albatrosses (Croxall. et al. 1992) or Common Terns (Gonzaléz-Solís, Becker, Jover & Ruiz, submitted ms). Except higher recruiting probabilities of Short-tailed Shearwaters reared by first breeders there is little evidence for strong effects of parental age on recruitment. Longer-lived individual Short-tailed Shearwaters were more productive than shorter-lived individuals, especially in their early breeding years (Wooller et al. 1990), and high offspring production during the breeding career was associated with a decreased mortality rate (Bradley et al. 1989). These facts again indicate positive quality effects on both fledgling production as well as on survival. In the Common Tern age played a negligible role with respect to body condition (Wendeln & Becker in press) and with respect to the number of young prospecting, in contrast to other factors (Table 4 and Table 5).
Quality
Quality of a breeder has been described by different parameters, including the ability to produce large eggs (Bolton 1991) or clutches (Coulson & Porter 1985), the ability to breed early (Mills 1989, Sydeman & Eddy 1995), the amount of food provided (e.g. Nisbet 1973) or the body condition (Weimerskirch 1992, Wendeln & Becker in press). Also longevity is related to adult quality: Good quality Kittiwakes characterised by consistent high reproductive success also live longer (Coulson & Thomas 1985). Individual Short-tailed Shearwaters that lived for longer had also a higher average reproductive success (Wooller et al. 1990). Weimerskirch (1992) explaines the cross-sectional increase of body condition, egg size (see also Croxall et al. 1992) and breeding success with age of the Wandering Albatross Diomedea exulans by progressive selection of high quality birds surviving longer than ‘bottom’ birds.
In the Common Tern, our data show that individual body condition is a consistent quality trait over many years increasing only at young age if at all. Birds in good condition allocated more effort to offspring. Consequently the individual condition correlates positively with chick growth as well as with reproductive success (Wendeln & Becker in press). From that we could expect that in female Common Tern mass may be also directly related to the survival of chicks to recruitment, what is substantiated by the initial data presented here. In the Red-billed Gull, body mass was not related to recruitment; it decreased, however, with increasing laying date (Mills 1989). Early laying is found to be another indicator of adult quality in many seabird species investigated (see above).
Common Terns rearing more chicks to fledging per season can ‘expect’ to have also more prospecting and recruiting progeny. Selection should favour those adults which have the quality to perform in this way. As breeding success and condition is consistent over years (Wendeln & Becker in press) we suppose that high quality adult Common Terns will also live longer, like Thomas & Coulson (1988) described in Kittiwakes. Thus good conditional state seems to enhance recruitment through both high annual breeding success and longevity. That mates of similar condition are paired (Wendeln et al. 1997) may strengthen the positive effects of condition on fitness, and may be also subject to sexual selection.
Condition seems to be one important parameter of quality. What constitutes condition? Besides e.g. phenotypic traits like physiological state, skeleton and muscle characteristics or nutritional reserves (see review McNamara & Houston 1996), foraging competence will be of great importance, especially in seabirds. Foraging decisions are linked with adult body mass and brooding allocation (Weimerskirch 1998). Individuality of foraging patterns as well as learning to exploit prey is well known in marine birds and could account for their delayed age of commencement of breeding (Hunt et al. 1999). Inter-individual variation in cognitive performances could cause differences in foraging abilities and skills to use marine food sources efficiently, and should be considered as a possible component of an individual’s conditional state.
Heritability
About three quarters of the differences between individual Manx Shearwaters in body size are due to genetic causes (Brooke 1990). Like morphological parameters also the quality of seabirds is a very consistent individual trait maintained for lifetime (e.g. Coulson & Porter 1985, Porter & Coulson 1988, Cobley et al. 1998); hence, Coulson & Porter (1985) assume that it may well have a genetical component. In the Common Tern body condition is stable over years (Wendeln & Becker in press), but heritability of condition remains to be proved.
There are many data showing that reproductive traits like clutch and egg size are of high repeatability in birds, also in seabirds (Coulson & Thomas 1985; Thomas & Coulson 1988; Mills 1989; Croxall et al. 1992; Weimerskirch 1992; Cobley et al. 1998; Wendeln & Becker in press; Gonzaléz-Solís, Becker, Jover & Ruiz, unpubl. ms). In birds, traits related to reproductive performance have significant but low heritabilities in the range of 30 – 40% (Boag & van Nordwijk 1987). In seabirds, however, heritability studies including across generations changes with respect to reproductive performance are rare owing to logistical problems. Brooke (1986) described high heritability of fledging age in Manx Shearwaters. Mills (1989) could compare reproductive characteristics of Red-billed Gull parents and offspring (five breeding seasons for each generation). No significant correlations emerged between the number of chicks fledged by male or female parents and that fledged by their offspring. Also in the number of recruits, in frequency and onset of breeding no significant intergenerational relationships were found. Neither age nor area of first breeding is transmitted from Kittiwake parents to offspring (Wooller & Coulson 1977), suggesting that these traits may be acquired during a birds ontogenetic history.
CONCLUSIONS
What can we learn from the information on parental quality and recruitment in seabirds compiled in this paper? The close relationship between breeding life-span and number of reproducing offspring underlines the importance of longevity on which strong selection pressures may act in seabirds. Longevity seems to be related to high individual quality and body condition, manifested also in an early seasonal onset of laying, which affects fledglings also positively in a direct way. Species- and age-specific trade-offs between survival and fecundity lead to an increase in progeny production during breeding life. In species rearing more than one chick, annual fledging success is maximised to enhance recruiting probability of offspring, also by direct positive effects on survival of fledglings. Improvement of food provisioning of the young by the parents increase post-fledging survival, competitive abilities of young and facilitate recruitment in young age. These outlined characteristics shape seabird life-styles (e.g. Furness & Monaghan 1987), across several systematically different groups, and seem to be of adaptive significance for long-lived birds in the marine environment. Understanding the determinants of variation in individual quality is an important task for future studies.
ACKNOWLEDGEMENTS I thank F. Bairlein, J. Gonzaléz-Solís, P. Monaghan and H. Wendeln for their comments improving this paper. I´m grateful to all my colleagues who helped with field work and data analyses in the Common Tern project, which was supported by the Deutsche Forschungsgemeinschaft (Be 916/3).REFERENCES
Becker, P.H. 1996. Flußseeschwalben (Sterna hirundo) in Wilhelmshaven. Oldenb. Jahrbuch 96: 263-296.
Becker, P.H. & Wendeln, H. 1997. A new application for transponders in population ecology of the Common Tern. Condor 99: 534-538.
Boag, P.T. & van Nordwijk, A.J. 1987. Quantitative genetics. In: Cooke, F. and Buckley, P.A. (eds.) Avian genetics. New York: Academic Press: 45-78.
Bolton, M. 1991. Determinants of chick survival in the Lesser Black-backed Gull: Relative contribution of egg size and parental quality. J. Anim. Ecol. 60: 949-960.
Boulinier, T., Danchin, E., Monnat, J.-V., Doutrelant, C. & Cadiou, B. 1996. Timing of prospecting and the value of information in a colonial breeding bird. J. Avian Biology 27: 252-256.
Bradley, J.S., Skira, I.J. & Wooller, R.D. 1990. A long-term study of Short-tailed Shearwaters Puffinus tenuirostris on Fisher Island, Australia. Ibis 133 suppl. I: 55-61.
Bradley, J.S. & Wooller, R.D. 1991. Philopatry and age of first-breeding in long-lived birds. Acta XX Congr. Intern. Ornith. 1657-1665.
Bradley, J.S., Wooller, R.D., Skira, I.J. and Serventy, D.L. 1989. Age-dependent survival of breeding Short-tailed Shearwaters Puffinus tenuirostris. J. Anim. Ecol. 58: 175-188.
Brooke, M. 1986. Manx Shearwater chicks: seasonal, parental, and genetic influences on the chick’s age and weight at fledging. Condor 88: 324-327.
Brooke, M. 1990. The Manx Shearwater. Poyser, London.
Burger, J. 1980. The transition to independence and postfledging parental care in seabirds. In: Burger, J., Olla, B.L. and Winn, H.E. (eds) Behavior of marine animals. Vol. 4; marine birds; New York; Plenum: 367-447.
Clutton-Brock, T.H. 1988. Reproductive success. Chicago. The University of Chicago Press.
Cobley, N.D., Croxall, J.P. & Prince, P.A. 1998. Individual quality and reproductive performance in the Grey-headed Albatross Diomedea chrysostoma. Ibis 140: 315-322.
Cooke, F., Rockwell, R. F. & Lank, D.B. 1991. Recruitment in long-lived birds: genetic considerations. Acta XX Congr. Intern. Ornith. 1666-1677.
Coulson, J.C. & Porter, J.M. 1985. Reproductive success of the Kittiwake Rissa tridactyla: the roles of clutch size, chick growth rates and parental quality. Ibis 127: 450-466.
Coulson, J.C. & Thomas, C. 1985. Differences in the breeding performance of individual Kittiwake Gulls, Rissa tridactyla. In: Sibly, R. M. u. Smith, R. H. (eds.): Behav. Ecol.. Blackwell, Oxford, pp. 489-503.
Croxall, J.P., Rothery, P. & Crisp, A. 1992. The effect of maternal age and experience on egg-size and hatching success in Wandering Albatrosses Diomedea exulans. Ibis 134: 219-228.
Danchin, E., Cadiou, B., Monnat, J. Y. & Estrella, R. R. 1991. Recruitment in long-lived birds: conceptual framework and behavioural mechanisms. Acta XX Congr. Intern. Ornith. 1641-1655.
Dann, P. & Cullen, J. M. 1990. Survival, patterns of reproduction, and lifetime reproductive output in Little Blue Penguins (Eudyptula minor) on Phillip Island, Victoria, Australia. In: Davis, L. S. u. Darby, J. T. (eds.): Penguin Biology. Academic Press, Inc., New York, pp. 63-84.
Forslund, P. & Pärt, T. 1995. Age and reproduction in birds - hypotheses and tests. TREE 10: 374-378.
Furness, R.W. & Monaghan, P. 1987. Seabird Ecology. Blackie, Glasgow and London.
Gaston, J.A.J. 1985. Development of the young in the Atlantic Alcidae. In: Nettleship, D.N. & Birkhead, T.R. (eds.) The Atlantic Alcidae. Academic Press, London.
Halley, D.J., Harris, M.P. & Wanless, S. 1995. Colony attendance patterns and recruitment in immature Common Murres (Uria aalge). Auk 112: 947-957.
Harris, M.P., Halley, D. J. & Wanless, S. 1992. The post-fledging survival of young Guillemots Uria aalge in relation to hatching date and growth. Ibis 134: 335-339.
Harris, M.P. & Rothery, P. 1985. The post-fledging survival of young Puffins Fratercula arctica in relation to hatching date and growth. Ibis 127: 243-250.
Hedgren, S. 1981. Effects of fledging weight and time of fledging on surval of Guillemots Uria aalge chicks. Ornis Scand. 12: 51-54.
Hunt, G.L., Mehlum, F., Russell, R.R., Irons, D., Decker, M.B. & Becker, P.H. 1999. Physical processes, prey abundance, and the foraging ecology of seabirds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2040-2056. Johannesburg: BirdLife South Africa.
Jarvis, M.J.F. 1974. The ecological significance of clutch size in the South African Gannet (Sula capensis [Lichtenstein]). J. Anim. Ecol. 43: 302-352.
Lloyd, C.S. 1979. Factors affecting breeding of Razorbills Alca torda on Skokholm. Ibis 121: 165-176.
McNamara, J.M. & Houston, A.I. 1996. State-dependent life histories. Nature 380: 215-221.
Mills, J.A. 1989. Red-billed Gull. In: Newton, I. (ed.) I. Lifetime reproduction in birds. Academic Press, London, pp. 387-404.
Newton, I. 1989. Lifetime reproductive success in birds. London, Academic Press.
Nisbet, I.C.T. 1973. Courtship-feeding, egg size and breeding success in Common Terns Sterna hirundo. Nature 241: 169-178.
Nisbet, I.C.T. 1996. Post-fledging survival in Common Terns in relation to brood order, hatching date and parental age. Colonial Waterbirds 19: 253-255.
Nisbet, I.C.T. & Drury, W.H. 1972. Post-fledging survival in Herring Gulls in relation to brood-size and date of hatching. Bird-Banding 43: 161-240.
Nisbet, I.C.T., Spendelow, J.A. & Hatfield, J.S. 1995. Variation in growth of Roseate Tern chicks. Condor 97: 335-344.
Nisbet, I.C.T., Winchell, J.M. & Heise, A.E. 1984. Influence of age on the breeding biology of Common Terns. Colonial Waterbirds 7: 117-126.
Ollason, J.C. & Dunnet, G.M. 1978. Age, experience and other factors affecting the breeding success of the Fulmar, Fulmarus glacialis, in Orkney. J. Anim. Ecol. 47: 961-976.
Ollason, J.C. & Dunnet, G.M. 1988. Variation in breeding success in Fulmars. In: Clutton-Brock, T. H. (ed.) Reproductive success: Studies of individual variation in contrasting breeding systems. The University of Chicago Press, Chicago, London, pp. 263-278.
Parsons, J., Chabrzyk, G. & Duncan, N. 1976. Effects of hatching date on post-fledging survival in Herring Gulls. J. Anim. Ecol. 45: 667-675.
Perrins, C.M., Harris, M.P. & Britton, C.K. 1973. Survival of Manx Shearwaters Puffinus puffinus. Ibis 115: 535-548.
Porter, J. M. 1988. Prerequisites for recruitment of Kittiwakes Rissa tridactyla. Ibis 130: 204-215.
Porter, J. M. & Coulson, J. C. 1997. Long-term changes in recruitment to the breeding group, and the quality of recruits at a Kittiwake Rissa tridactyla colony. J. Anim. Ecol. 56: 675-690.
Price, T.D. & Boag, P.T. 1987. Selection in natural bird populations. In: Cooke, F. and Buckley, P.A. (eds.) Avian genetics. New York: Academic Press: 257-287.
Rattiste, K. & Lilleleht, V. 1986. Some aspects of the demography of the Common Gull Larus canus in Estonia. Var Fagelv. Suppl. II: 179-186.
Spear, L. & Nur, N. 1994. Brood size, hatching order and hatching date: effects on four life-history stages from hatching to recruitment in Western Gulls. J. Anim. Ecol. 63: 283-298.
Spendelow, J. A. 1991. Postfledging survival and recruitment of known-origin Roseate Terns (Sterna dougallii) at Falkner Island, Connecticut. Colonial Waterbirds 14: 108-115.
Sydeman, W.J. & Eddy, J.O. 1995. Repeatability in laying date and its relationship to individual quality for Common Murres. Condor 97: 1048-1052.
Thomas, C.S. & Coulson, J.C. 1988. Reproductive success of Kittiwake Gulls, Rissa tridactyla. In: Clutton-Brock, T. H. (ed.) Reproductive success: Studies of individual variation in contrasting breeding systems. The University of Chicago Press, Chicago, London, pp. 251-262.
Viksne, J. & Janaus, M. 1993. Post-fledging survival of the Black-headed Gull (Larus ridibundus) depending on hatching parameters. Ring 15: 387-389.
Weimerskirch, H. 1992. Reproductive effort in long-lived birds: Age-specific patterns of condition, reproduction and survival in the Wandering Albatross. Oikos 64: 464-473.
Weimerskirch, H. 1998. How can a pelagic seabird provision ist chick when relying on a distant food source? Cyclic attendance at the colony, foraging decision and body condition in sooty shearwaters. J. Anim. Ecol. 67: 99-109.
Wendeln, H. 1997a. Body mass of female Common Terns (Sterna hirundo) during courtship: relationships to male quality, egg mass, diet, laying date and age. Colonial Waterbirds 20: 235-243.
Wendeln, H. 1997b. Allocation of parental duties and foraging behaviour influence body condition of adult Common Terns, Sterna hirundo. Bird Behaviour 12: 47-54.
Wendeln, H. & Becker, P.H. 1996. Body mass change in breeding Common Terns (Sterna hirundo). Bird Study 43: 85-95.
Wendeln, H. & Becker, P.H. 1998. Populationsbiologische Untersuchungen an einer Kolonie der Flußseeschwalbe (Sterna hirundo). Vogelwelt 119: 209-213.
Wendeln, H. & Becker, P.H. In press. Effects of parental quality and effort on the reproduction of Common Terns Sterna hirundo. J. Anim. Ecol.: In press.
Wendeln, H., Becker, P.H. & Wagener, M. 1997. Beziehungen zwischen Körpermasse und Körpergröße bei Paarpartnern der Flußseeschwalbe Sterna hirundo. Vogelwarte 39: 141-148.
Wiggins, D.A., & Morris, R.D. 1987. Parental care in the Common Tern Sterna hirundo. Ibis 129: 533-540.
Wooller, R.D., Bradley, J.S., Skira, I.J. & Serventy, D.L. 1989. Short-tailed Shearwater. In: Newton, I. (ed.) Lifetime reproduction in birds. Academic Press.,
Wooller, R.D., Bradley, J.S., Skira, I.J. & Serventy, D.L. 1990. Reproductive success of Short-tailed Shearwaters Puffinus tenuirostris in relation to their age and breeding experience. J. Anim. Ecol. 59: 161-170.
Wooller, R.D. & Coulson, J.C. 1977. Factors affecting the age of first breeding of the Kittiwake Rissa tridactyla. Ibis 119: 339-349.
Table 1. Proportion of young Common Terns recorded as natal prospectors at the colony site in Wilhelmshaven. Year classes 1992-1995 combined, n fledglings in brackets. No significant differences by number of fledglings (c 2-test, c 2 = 0.75, P = 0.69) or hatching order (c 2 = 1.52, P = 0.47).
Table 2. Relationships of age of recruitment of Common Terns and date of first return to the natal colony as prospector, chick peak mass and prospector mass.

Table 3. Spearman-rank correlations between average numbers of fledglings, prospectors and recruits produced by Common Tern males (n=38) or females (n=42) per year (only birds with at least one fledgling, data from 1992-1994; prospecting or recruiting until 1997). As more prospectors are expected to recruit after 1997, correlations with recruits will be minimum figures. * p < 0.05, ** p<0.01, *** p < 0.001.

Table 4. Relationships between the number of natal prospectors produced by individual Common Terns and adult age, body mass during incubation and reproductive performance. Average values of birds studied at least for three years from 1992-1995. Spearman correlation coefficients.
Table 5. Effects of individual parental traits on recruitment and of natal characteristics influencing survival of fledglings to recruitment. +, -: positive, negative effects; (+), (-): weak effects, or present only in some years; 0: no effect.

1
Wooller et al. (1989); 2 Wooller et al (1990); 3 Mills (1989); 4 this paper, preliminary data, until prospection; 5 Perrins (1966); 6 Perrins et al. (1973); 7 Jarvis (1974); 8 Nisbet & Drury (1972); 9 Parsons et al. (1976); 10 Spear & Nur (1994), original brood size; 11 Viksne & Janaus (1993); 12 Wooller & Coulson (1977); 13 Coulson & Porter (1985); 14 Nisbet (1996); 15 Hedgren (1981); 16Harris et al. (1992); 17 Harris & Rothery (1985); 18 Lloyd (1979)
Fig. 1. Correlation of chick peak mass with prospector body mass in the year of first prospection at the natal colony in Common Tern immatures from Wilhelmshaven
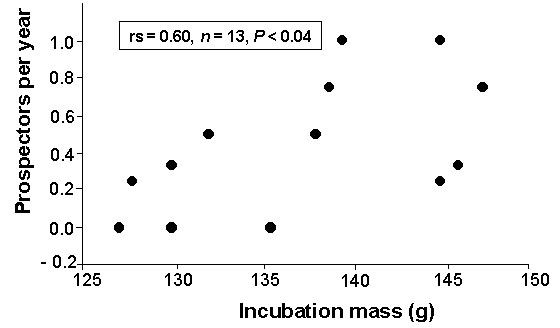
Fig. 2. Relationship between female Common Tern body mass during incubation and their number of prospectors per year. Average values for females studied 1992-1995 at least for three breeding seasons.