S24.3: Ecological factors affecting hunting behaviour during the post-fledging dependence period of raptors
Javier Bustamante
Department of Applied Biology, Estación Biológica de Doñana, CSIC. Avda. María Luisa s/n. 41013- Sevilla, Spain,
fax 34 95 4621125, e-mail busta@ebd03.ebd.csic.es Bustamante, J. 1999. Ecological factors affecting hunting behaviour during the post-fledging dependence period of raptors. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1381-1396. Johannesburg: BirdLife South Africa.The post-fledging dependence period is the time after leaving the nest during which raptor fledglings depend on their parents for food. Fledglings of some species seem to acquire independence without having hunted by themselves. In other species hunting behaviour matures gradually during this period. First prey captured by fledglings are frequently invertebrates, even if these are not an habitual prey of the species. Play with objects, and social play are frequent among fledglings during this period. They are more common in species hunting agile prey (birds, mammals) and in those with more elaborate hunting techniques. This agrees with predictions of the hypothesis that relate play behaviour with the maturation of the ability to capture and manipulate prey, and the training of the muscles necessary for these activities. Experimental work with the American Kestrel has shown that fledglings prefer to play with objects that resemble their habitual prey. Hunting behaviour and prey recognition seem to be innate in raptors and develop in fledglings even if they cannot observe other individuals. In some species there can be some social facilitation, so that successful hunting techniques or places to search for prey are learnt by observing other individuals.
INTRODUCTION
The post-fledging dependence period is the time after leaving the nest in which fledglings are still dependent on their parents for food and protection. In raptors the duration of this period can range from 5 days in the Lesser Kestrel Falco naumanni (Bustamante & Negro 1994) to almost one year in the Crowned Eagle Stephanoaetus coronatus (Brown 1966). At the time of fledging growth is almost complete. Raptor fledglings have adult body size and only flight feathers continue to grow for some days or weeks. During the post-fledging dependence period fledglings learn how to fly and train the muscles necessary for this activity. They probably also learn or start to develop the behavioural repertoire necessary for independent life. Raptors, particularly those feeding on live prey, require elaborated hunting techniques to obtain their food. Maturation of the hunting techniques of young raptors could take place during the post-fledging dependence period.
There are few detailed studies on the maturation of hunting behaviour in raptors. Meinertzhagen (1954) reported how young Ospreys Pandion haliaetus learned to fish by observing their parents, but Schaadt and Rymon (1982) demonstrated that fishing behaviour is innate in Ospreys, and that they actually do not need to watch other Ospreys to learn. Information on how young raptors learn to hunt comes mainly from observational studies during the post-fledging dependence period (see e.g. Johnson 1986; Bustamante 1993a; 1993b), and anecdotal observations (see e.g. Cade 1953; Simmons 1984), but very few experimental studies (Negro et al. 1996). Young raptors and occasionally adults have been observed playing with objects (Bildstein 1980; Paull 1981; Romer 1987). This behaviour has been interpreted as a mechanism to acquire the ability to capture and manipulate prey (Ficken 1977) or to train the muscles associated with these activities (Fagen 1981).
I have made a review of the literature on hunting behaviour of raptors during the post-fledging dependence period and reanalysed my own data to find some general patterns on the maturation of hunting behaviour. I have tried to answer the following questions: Do young raptors capture prey by themselves before becoming independent? Is hunting behaviour innate or has it to be learned? Do young raptors learn to recognise prey, hunting habitats or hunting behaviour by observing other individuals? Is there some kind of ‘teaching’ of hunting behaviour by parents? Which is the role of object and social play in the maturation of hunting behaviour?
METHODS
I have reviewed the literature on the post-fledging dependence period of raptors plus unpublished observations that give some information on behaviour of raptor fledglings during this time. Studies are very heterogeneous in quality and range from very detailed studies with numerous radio-tagged individuals observed for several hundred hours to opportunistic or anecdotal observations of one individual. I have kept information coming from different sources separated in tables. Considering the information available for each species I have tried to distinguish between studies that document the absence of certain behaviours (e.g. play with objects, successful hunting) --provided that study the was detailed enough-- and studies that allow no inference on whether a certain behaviour could take place or not during the post-fledging dependence period. Even though, the lack of observations does not necessarily mean that certain behaviour does not take place in that species.
HUNTING DURING THE POST-FLEDGING DEPENDENCE PERIOD
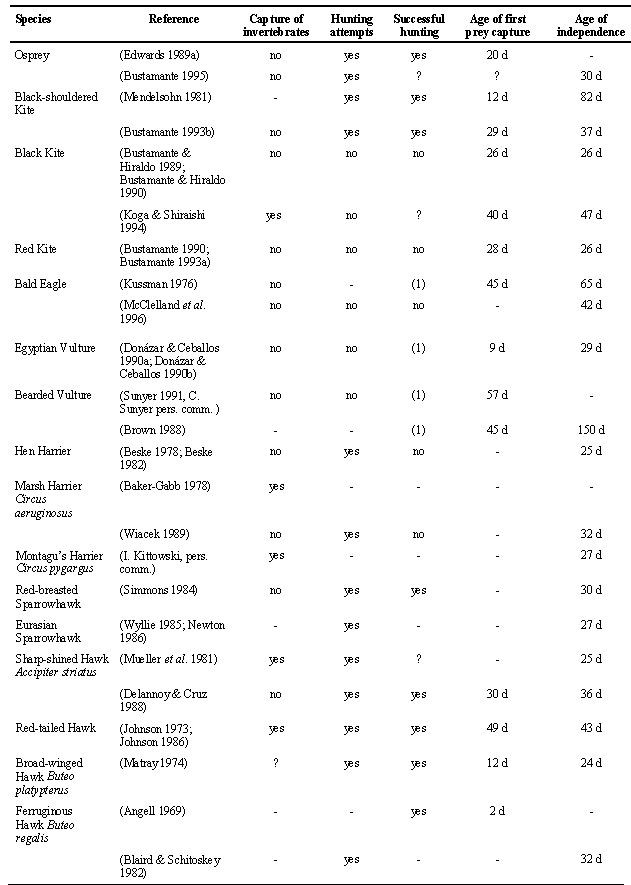
In some species of raptors fledglings become independent from their parents without having made any prey capture by themselves. That is, independence from parents and first hunting attempts seem to take place almost at the same time and there is not an apparent gradual maturation of hunting or searching behaviour during the post-fledging dependence period (Table 1). This behaviour is common in carrion feeders like milvus kites Milvus spp., the Bald Eagle Haliaeetus leucocephalus, and vultures (Bearded vulture Gypaetus barbatus, Egyptian vulture Neophron percnopterus). Hunting attempts during post-fledging have rarely been observed in carrion feeders. Fledglings have been observed in some instances feeding on carcasses before becoming independent (50% of studies, n = 8), but no attempts to capture living prey have been recorded and only rarely have they been observed capturing invertebrates (14% of studies, n = 7). In detailed studies with kites (Red Kite Milvus milvus and Black Kite M. migrans) the mean age at which kites were captured at baited traps was the same as the mean age of independence from parents (Table 1). Fledgling kites still dependent on their parents for food were never observed trying to hunt nor captured in baited traps (Bustamante & Hiraldo 1989; Bustamante 1990; Bustamante 1993a).
The hunting behaviour of raptor species feeding on more active prey mature gradually before independence (Table 1). Hunting attempts by fledglings still dependent on their parents for food have been observed in Ospreys, Black-shouldered Kites Elanus caeruleus, harriers Circus spp., accipiter hawks Accipiter spp., buzzards Buteo spp. eagles (Aquila spp., Hieraeetus spp., Crowned Eagle) and falcons Falco spp. In most species fledglings start capturing invertebrates, even if these are not the habitual prey of the species. Capture of invertebrates before independence is very common in falcons. It has been reported in 9 out of 10 species, and in 94% of the studies of post-fledging behaviour of falcons (n = 16). The only exception in falcons seems to be the Lesser Kestrel that feeds on invertebrates. Fledglings were not observed capturing invertebrates probably because of the very short post-fledging dependence period (5 days). Invertebrate capture is also common in buzzards (75% of studies, n = 4), harriers (40%, n = 5) and accipiter hawks (33%, n = 3). It has not been observed in eagles (n = 6) nor in specialised hunters like the Osprey or the Black-shouldered Kite.
Fledglings of raptors with specialised feeding habits seem to be able to hunt typical adult prey successfully before independence (Table 1). Fledgling Ospreys capture fish (Edwards 1989a; Bustamante 1995) and Black-shouldered Kites capture mice (Mendelsohn 1981; Bustamante 1993b) while still dependent on their parents for food. Successful hunting of prey other than invertebrates has been observed in buzzards (80% of studies, n = 5), falcons (43%, n = 14) , eagles (43%, n = 7), accipiter hawks (one study) but not in harriers (n = 2).
There are very few studies that have measured changes in hunting success of young raptors during the post-fledging dependence period. Improvement in hunting success with age has been documented for the Osprey (Edwards 1989b), the American Kestrel Falco sparverius (Varland et al. 1991; Varland & Loughin 1992) and the Greater Kestrel Falco rupicoloides (McCann & Kemp 1994).
IS HUNTING BEHAVIOUR INNATE OR HAS TO BE LEARNED?
Although Meinertzhagen (1954) described how Ospreys taught their offspring how to fish, the work by Schaadt and Rymon (1982) shows that fishing behaviour is innate in Ospreys. Schaadt and Rymon showed that a hand reared Osprey was able to recognise the habitual prey of the species and learn how to fish without any previous observation of an Osprey fishing. It has been established that even an elaborated behaviour like using stones as tools to break eggs, is innate in the Egyptian Vulture Neophron percnopterus (Thouless et al. 1989) and in the Black-breasted Buzzard Hamirostra melanosternum (Debus 1991). Inexperienced juveniles do not have to observe other individuals breaking eggs to learn, and there is no evidence of cultural transmission in this behaviour (Thouless et al. 1989).
The detailed work by Edwards (1989b; 1989a) about fishing progression in juvenile Ospreys, indicates that juveniles do not normally learn by imitation of adults (they do not follow their parents to the places where they fish). Even though Edwards (1989b) observed that siblings that remained together learned faster than singletons, and that individuals that learn together imitate those fishing techniques that are successful. In that way that siblings develop similar fishing techniques and similar fish preferences. In other species there could also be some kind of social facilitation (Thorpe 1963) in the development of hunting behaviour. Juvenile American Kestrels tend to join together in groups during and after the post-fledging dependence period and hunt socially (Varland et al. 1991). It has been suggested that there could be some imitative learning during this period although it has not been proved that big broods learn faster than small broods (Varland & Loughin 1992).
If raptor fledglings learned hunting techniques imitating their parents it would be necessary that offspring followed their parents to their hunting grounds. Although this has been described in some species, the contrary has also been observed at least in half of the species studied (Table 2). To follow the parents seems to be relatively common in vultures, both in new-world vultures (Wallace & Temple 1983; Rabenold 1986) and in old-world vultures (Brown 1988; Heredia & Sunyer 1989; Donázar & Ceballos 1990b), but seems to be uncommon in other species of raptors. It has only been observed by Lowe (1978) and Johnson (1986) in the Red-tailed Hawk Buteo jamaicensis, by Bustamante (1993b) in the Black-shouldered Kite and by Ratcliffe (1980) in the Peregrine Falcon Falco peregrinus. Following adults to carcasses is a behaviour that could be expected in vultures because these species rely to a great extent on observing the behaviour of other individuals or other raptors to locate carcasses. In other species, following the parents to the hunting area seems to happen only when the hunting area is small and is located far from the nest as was the case in one Black-shouldered Kite pair I studied (Bustamante 1993b).
Apart from the paper of Meinertzhagen (1954) there are few observations that suggest some kind of active teaching of hunting techniques to offspring. Air transfer of prey from parent to offspring during the post-fledging dependence period could be a mechanism evolved to facilitate the training of offspring in the capture of agile prey. If this was so it could be considered some kind of active training or teaching of hunting techniques by adults.
During the post-fledging dependence period the most common prey transfer from parents to offspring takes place with one or both individuals perched. Air transfers are not uncommon in some species and have been observed in the Black-shouldered Kite, harriers, accipiter hawks, buzzards, eagles, and falcons (Table 2). The most common air transfer is the talon-to-talon, or talon-to-beak transfer of prey in mid-air and has been recorded at least in 25 species. In some species parents drop the dead prey and offspring recapture it in mid air or on the ground. These transfers are called ‘dead-drops’ and have been recorded in 11 species. The most elaborate air transfer is the one called ‘live-drop’ in which a live prey is released by a parent and left to the offspring to be recaptured. It has been observed only in the Northern Goshawk Accipiter gentilis (Palmer 1988), Eurasian Sparrowhawk Accipiter nisus (Newton 1986), Cooper´s Hawk Accipiter cooperi (H. McElroy in Spofford & Amadon 1993), New Zealand Falcon Falco novaeseelandiae (Lawrence & Gay 1991) and Peregrine Falcon (Sherrod 1983). According to Newton (1979; 1986) ‘live-drops’ in accipiter hawks would not require a special modification of adult behaviour. It is frequent that the hawk does not kill the prey during capture so a ‘live-drop’ would not substantially differ from a ‘dead-drop’ that is very common in this group. ‘Live-drops’ in falcons are a more elaborate behaviour because it requires that the falcon does not kill its prey immediately after capture as it normally does (Newton 1979; Sherrod 1983).
Air prey transfers during the post-fledging dependence period could have other functions. It has been suggested that they could be a mechanism used by parents to test the flying and prey capture capacity of offspring, and according to this decide when to stop parental investment (Simmons 1984; Ferrer 1990). In species with a prolonged post-fledging dependence period, they could be a mechanism of brood reduction in case of an unexpected decline in food availability during the post-fledging dependence period, because it is only the first fledgling to approach the parent that receives the prey. Also it could be a fast way to identify and feed the hungriest fledgling in a multiple brood and serve a similar function as feeding chases in pygoscelid penguins (Bustamante et al. 1992).
Species performing air prey transfers have in common that they feed on agile prey like birds and small mammals, tend to have large broods (three or more chicks), and that air prey transfers are also performed between breeding adults. It is possible that some kinds of prey are not adequate for air transfers. This would be the case in species feeding on large prey (e.g. Crowned Eagle, Black Eagle Aquila verreauxi), slippery prey like fish (e.g. Osprey, Bald Eagle) or prey difficult to transport or manipulate like carrion or bones (e.g. Egyptian Vulture, Bearded Vulture). Air prey transfer requires an extra energy expenditure for fledglings and this could explain why they do not take place with small prey like insects (e.g. Lesser Kestrel ).
The information available suggest that prey recognition and probably hunting techniques are innate in raptors. There can be some learning by imitation, probably more from other juveniles than from adults. Air prey transfers could have some role in maturation of hunting behaviour in some raptor species, probably similar to object and social play (as it will be seen below), but it is difficult to evaluate its importance. The anecdotal occurrence of the more elaborate ‘life-drops’ suggest that an active teaching by adult is not very important in the maturation of hunting behaviour of juvenile raptors.
PLAY AND ITS RELATION WITH MATURATION OF HUNTING BEHAVIOUR
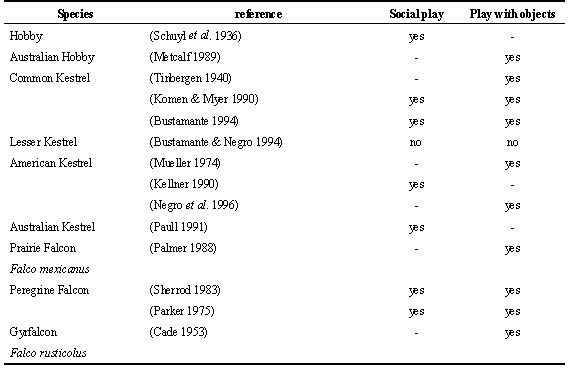
Play seems to have an important function in the maturation of hunting behaviour as well as in the maturation of flight during the post-fledging dependence period. According to Ficken (1977) play could be an important way to acquire ability to capture and manipulate prey or even train the muscles associated with these behaviours (Fagen 1976). Different kinds of play behaviour have been described in birds of prey, mainly in juveniles, but also in adults (see e.g. Bildstein 1980; Paull 1981). In general, play behaviour in raptors can be classified as ‘object manipulation play’, which consists in the practice upon inanimate objects of different behaviours related with prey capture, prey transport or prey consumption, and ‘social play’ in which juveniles chase in a playful way mainly their siblings, but also other juveniles of the same species, other raptors, other birds, or mammals in fictitious attacks, apparently without any intention of making a capture or hurting the other individual. Social play or chases have been observed in aprox. 84% of studies of post-fledging behaviour in raptors and at least in 26 species. Object play has been described in approx. 74% of studies and in 25 species (Table 3). It is difficult to compare data from different authors on different species because there are many factors that influence the observation of play behaviour among juveniles. Also, chases among juveniles or towards other species are not always considered social play and have received different interpretations like: unsuccessful hunting attempts, competition among siblings, or nest defence against intruders or predators (Rosenfield & Kanvik 1983; Hiraldo et al. 1989; Donázar & Ceballos 1990b).
Object play has been observed in all kind of raptors but seems to be more frequent in those species with elaborate hunting techniques or feeding on more agile and difficult to capture prey like small mammals, birds or fish (i.e. Common Kestrel Falco tinnunculus, Black-shouldered kite, Osprey or Peregrine Falcon) and relatively uncommon in those species with generalist feeding habits like the milvus kites (Bustamante & Hiraldo 1989; Bustamante 1993a; Koga & Shiraishi 1994) or the Bald Eagle (Kussman 1976), feeding on carrion like vultures (Donázar & Ceballos 1990a; Donázar & Ceballos 1990b) or on insects like the Lesser Kestrel (Bustamante & Negro 1994). It is odd that object play has not been recorded more often in specialised bird feeders like accipiter hawks (see e.g. Newton 1979; Wyllie 1985; Newton 1986; Palmer 1988; Kenward et al. 1993a; 1993b; Frumkin 1994). Being forest raptors this is probably due to the difficulty of observing the behaviour of these species at close range. In fact, when accipiter hawks have been observed at close range or in captivity, play with objects has been documented as frequent (Simmons 1984; Dewhurst 1986; Aumann 1988).
Chases and social play among fledglings that could be more related with the maturation of flight behaviour seem to be more frequent than object play in most species, and some times they are almost the only play behaviour recorded in species with generalised feeding habits (i.e. milvus kites or vultures).
Play in birds of prey, in any case, does not seem to have such an important function for learning as in mammals (Ficken 1977). It has been suggested that the frequency in which play is recorded in raptor fledglings is inversely proportional to the abundance of real prey with which to practice (Sherrod 1983). An experiment performed with captive American Kestrels corroborates this hypothesis (Negro et al. 1996). The experiment showed that kestrel fledglings devoted a fixed amount of time to object manipulation during the post-fledging dependence period. If offered the opportunity, they preferred to play with objects resembling their habitual prey but if these were absent play behaviour has redirected to other objects present in the pen. Bildstein (1980) observed that Hen Harriers Circus cyaneus chose to play with corn cobs of a size similar to their habitual prey. Also Red-breasted Sparrowhawks Accipiter rufiventris showed more interest in pine cones covered with crow feathers than in bare ones (Simmons 1984). The experimental work with American Kestrels also showed that there was some social facilitation and imitative behaviour in object play. Kestrels housed in the same pen tended to play with a similar intensity, preferred the same objects, and played more often on the same days. All this observations suggest that fledglings may be encouraged to play when they watch other individuals playing and indicates some role of social facilitation and imitative behaviour in learning of hunting techniques.
ACKNOWLEDGMENTS
I thank F. Hiraldo for suggestions during this work. Research funds during the writing of this review were provided by project PB96-0805 from DGES.
REFERENCES
Alonso, J.C., González, L.M., Heredia, B. & González, J.L. 1987. Parental care and the transition to independence of spanish imperial eagles (Aquila heliaca) in Doñana National Park, southwest Spain. Ibis 129: 212-224.
Angell, T. 1969. A study of the ferruginous hawk: adult and brood behavior. Living Bird 8: 225-241.
Aumann, T. 1988. Breeding behaviour of the brown goshawk Accipiter fasciatus. Australian Bird Watcher 12: 258-267.
Bahat, O. 1992. Post-fledging movements of golden eagles (Aquila chrysaetos homeyeri) in the Negev Desert, Israel, as determined by radio-telemetry. In: Priede, I.G. & Swift, S.M. (eds) Wildlife telemetry: Remote monitoring and tracking of animals. New York; Ellis Horwood: 612-621.
Baker-Gabb, D.J. 1978. Aspects of the biology of the Australasian harrier Circus aeruginous approximans. M. Sc. Thesis, University of Massey, Palmerstron, North New Zealand.
Beske, A.E. 1978. Harrier radio-tagging techniques and local and migratory movements of radio-tagged juvenile harriers. M.Sc. Thesis, University of Wisconsin, Stevens Point.
Beske, A.E. 1982. Local and migratory movements of radio-tagged juvenile harriers. Raptor Research 16: 39-53.
Bildstein, K.L. 1980. Corn cob manipulation in northern harriers. The Wilson Bulletin 92: 128-130.
Blaird, C.L. & Schitoskey, F. 1982. Breeding biology and diet of the ferruginous hawk in South Dakota. The Wilson Bulletin 94: 46-54.
Blumstein, D.T. 1990. An observation of social play in bearded vultures. Condor 92: 779-781.
Bollen, C. 1991. Breeding behaviour and diet of the Australian kestrel (Falco cenchroides). Australian Bird Watcher 14: 44-50.
Brown, C.J. 1988. A study of the bearded vulture Gypaetus barbatus in southern Africa. Ph.D. Thesis, University of Natal, Pietermaritzburg.
Brown, L.H. 1966. Observations on some Kenya eagles. Ibis 108: 531-572.
Bustamante, J. 1986. Conflicto paterno-filial e independencia de los pollos en el Milano negro. M.Sc. Thesis, University of Autónoma de Madrid, Madrid.
Bustamante, J. 1990. Condicionantes ecológicos del periodo de emancipación en Falconiformes. Ph. D. Thesis, University of Autónoma de Madrid, Madrid.
Bustamante, J. 1993a. Post-fledging dependence period and development of flight and hunting behaviour in the red kite Milvus milvus. Bird Study 40: 181-188.
Bustamante, J. 1993b. The post-fledging dependence period of the black-shouldered kite (Elanus caeruleus). Journal of Raptor Research 27: 185-190.
Bustamante, J. 1994. Behavior of colonial common kestrels (Falco tinnunculus) during the post-fledging dependence period in southwestern Spain. Journal of Raptor Research 28: 79-83.
Bustamante, J. 1995. The duration of the post-fledging dependence period of ospreys Pandion haliaetus at Loch Garten, Scotland. Bird Study 42: 31-36.
Bustamante, J., Cuervo, J. & Moreno, J. 1992. The function of feeding chases in the chinstrap penguin (Pygoscelis antarctica). Animal Behaviour 144: 161-166.
Bustamante, J. & Hiraldo, F. 1989. Post-fledging dependence period and maturation of flight skills in the Black Kite Milvus migrans. Bird Study 36: 199-204.
Bustamante, J. & Hiraldo, F. 1990. Factors influencing family rupture and parent-offspring conflict in the Black Kite Milvus migrans. Ibis 132: 58-67.
Bustamante, J. & Negro, J.J. 1994. The post-fledging dependence period of the lesser kestrel (Falco naumanni) in southwestern Spain. Journal of Raptor Research 28: 158-163.
Cade, T. 1953. Behavior of a young gyrfalcon. The Wilson Bulletin 65: 26-31.
Cramp, S. & Simmons, K.E.L. 1980. Handbook of the birds of Europe the Middle East and North Africa. Vol. II. Oxford; Oxford University Press:
Debus, S.J.S. 1984. Biology of the little eagle on the northern tablelands of New South Wales. Emu 84: 87-92.
Debus, S.J.S. 1991. Further observations on the black-breasted buzzard Hamirostra melanosternon using stones to break eggs. Australian Bird Watcher 14: 138-143.
Debus, S.J.S., Ley, A.J., Tremont, S. & Tremont, R. 1991. Breeding behaviour and diet of the australian hobby Falco longipennis in northern New South Wales. Australian Bird Watcher 14: 123-137.
Delannoy, C.A. & Cruz, A. 1988. Breeding biology of the Puerto Rican sharp-shinned hawk (Accipiter striatus venator). Auk 105: 649-662.
Dewhurst, C.F. 1986. The breeding ecology of the African goshawk at Karen, Nairobi, Kenya. Ostrich 57: 1-8.
Donázar, J.A. & Ceballos, O. 1990a. Acquisition of food by fledging Egyptian vultures (Neophron percnopterus) through nest-switching and acceptance by foster adults. Ibis 132: 603-617.
Donázar, J.A. & Ceballos, O. 1990b. Post-fledging dependence period and development of flight and foraging behaviour in the Egyptian vulture Neophron percnopterus. Ardea 78: 387-394.
Edwards, T.C., Jr. 1989a. The ontogeny of diet selection in fledgling ospreys. Ecology 70: 881-896.
Edwards, T.C., Jr. 1989b. Similarity in the development of foraging mechanics among sibling ospreys. Condor 91: 30-36.
Evans, S.A. 1977. Ecology and behavior of the Missisipi kite (Ictinia mississippiensis) in Southern Illinois. M. A. Thesis, University of Southern Illinois, Carbondale.
Fagen, R. 1981. Animal play behavior. New York; Oxford University Press:
Fagen, R.M. 1976. Exercise, play, and physical training in animals. In: Bateson, P.P.G. & Klopfer, P.H. (eds) Perspectives in Ethology. New York; Plenum: 189-219.
Ferrer, M. 1990. Dispersión juvenil en el águila imperial ibérica, Aquila adalberti. Ph. D. Thesis, University of Sevilla, Sevilla.
Ferrer, M. 1992. Regulation of the period of postfledging dependence in the Spanish imperial eagle Aquila adalberti. Ibis 134: 128-133.
Ficken, M.S. 1977. Avian play. Auk 94: 573-582.
Fitzner, R.E. 1978. Behavioral ecology of the Swainson's hawk Buteo swainsoni in southeastern Washington. Ph. D. Thesis, University of Washington State, Pullman.
Frumkin, R. 1994. Intraspecific brood-parasitism and dispersal in fledgling Sparrowhawks Accipiter nisus. Ibis 136: 426-433.
Gargett, V. 1972. Observations at a black eagle nest in the Matopos, Rhodesia. Ostrich 43: 77-108.
Heredia, R. & Sunyer, C. 1989. Seguimiento de la reproducción y dispersión juvenil del quebrantahuesos en los Pirineos, 1989. Memoria Interna ICONA.
Hiraldo, F., Delibes, M. & Estrella, R.R. 1989. Observations of a zone-tailed hawk family during the postfledging period. Journal of Raptor Research 23: 103-106.
Howard, R. & Powers, L. 1973. Hawks of the desert. Animal Kingdom 73: 22-27.
Ikeda, Y. 1985. Feeding by parents of golden eagle during early post-fledging period. Aquila chrysaetos 3: 15.
Johnson, S.J. 1973. Post-fledging activity of the red-tailed hawk. Raptor Research 7: 43-48.
Johnson, S.J. 1986. Development of hunting and self-sufficiency in juvenile red-tailed hawk (Buteo jamaicensis). Raptor Research 20: 29-34.
Kellner, C. 1990. Post-fledging behavior of American kestrels in Central Kentuky. Journal of Raptor Research 24: 56-58.
Kenward, R.E., Marcström, V. & Karlbom, M. 1993a. Post-nesting behaviour in goshawks, Accipiter gentilis: II. Sex differences in sociality and nest-switching. Animal Behaviour 46: 371-378.
Kenward, R.E., Marcström, V. & Karlbom, M. 1993b. Post-nestling behaviour in goshawks, Accipiter gentilis: I. The causes of dispersal. Animal Behaviour 46: 365-370.
Kochan, Z. 1987. Biologia orlika krzykliwego Aquila pomarina w okresie postpisklecym. Praca Magistèrska (M.Sc.) Thesis, University of Gdansk, Gdynia.
Koga, K. & Shiraishi, S. 1994. Parent-offspring relations during the post-fledging dependency period in the black kite (Milvus nigrans) in Japan. Journal of Raptor Research 28: 171-177.
Komen, J. & Myer, E. 1990. Observations on post-fledging dependence of kestrels (Falco tinnunculus rupicolus) in an urban environment. Journal of Raptor Research 23: 94-98.
Kussman, J.V. 1976. Post-fledging behavior of the Northern Bald Eagle, Haliaeetus leucocephalus alascanus Townsend, in the Chippewa National Forest Minnesota. Ph. D. Thesis, University of of Minnesota, Ann Arbor, Minnesota.
Laing, K. 1985. Food habits and breeding biology of merlins in Denali National Park, Alaska. Raptor Research 19: 42-51.
Lawrence, L.K. 1949. Notes on nesting pigeon hawks at Pimisi Bay, Ontario. The Wilson Bulletin 61: 15-25.
Lawrence, S.B. & Gay, C.G. 1991. Behaviour of fledgling New Zealand falcons (Falco novaeseelandiae). Notornis 38: 173-182.
Lowe, C.M. 1978. Certain life history aspects of red-tailed hawks in central Oklahoma and interior of Alaska. M. Sc. Thesis, University of Alaska, Fairbanks.
Matray, P.F. 1974. Broad-winged hawk nesting and ecology. Auk 91: 307-324.
McCann, K.I. & Kemp, A.C. 1994. Hunting behaviour of a fledgling greater kestrel Falco rupicoloides and its mother during the post-fledging period. Ostrich 1994: 1-6.
McClelland, B.R., McClelland, P.T., Yates, R.E., Caton, E.L. & McFadzen, M.E. 1996. Fledging and migration of juvenile bald eagles from Glacier National Park, Montana. Journal of Raptor Research 30: 79-89.
Meinertzhagen, R. 1954. The education of young ospreys. Ibis 96: 153-155.
Mendelsohn, J.M. 1981. A study of the blackshouldered kite Elanus caeruleus. Ph.D. Thesis, University of Natal, Pietermaritzburg.
Metcalf, E.C. 1989. The breeding biology of the Australian hobby Falco longipennis. Australian Bird Watcher 13: 20-29.
Mueller, H.C. 1974. The development of prey recognition and predatory behaviour in the American kestrel Falco sparverius. Behaviour 49: 313-324.
Mueller, H.C., Mueller, N.S. & Parker, P.G. 1981. Observation of a brood of sharp-shinned hawks in Ontario, with comments on the functions of sexual dimorphism. The Wilson Bulletin 93: 85-92.
Negro, J.J., Bustamante, J., Milward, J. & Bird, D.M. 1996. Captive fledgling American kestrels prefer to play with objects resembling natural prey. Animal Behaviour 52: 707-714.
Newton, I. 1979. Population ecology of raptors. Berkhamsted; T. & A.D.Poyser:
Newton, I. 1986. The sparrowhawk. Calton; T.& A.D.Poyser:
Oliphant, L.W. & Tessaro, S.V. 1985. Growth rates and food consumption of hand-raised merlins. Raptor Research 19: 79-84.
Palmer, N.G., Norton, P.M. & Robertson, A.S. 1985. Aspects of the biology of the forest buzzard. Ostrich 56: 67-73.
Palmer, R.S. 1988. Handbook of North American birds. Vol. 4. Diurnal raptors (Part 1). New York; Yale University Press:
Parker, A. 1975. Young male peregrines passing vegetation fragments to each other. British Birds 68: 242-243.
Parker, A. 1979. Peregrines at a Welsh coastal eyrie. British Birds 72: 104-114.
Paull, D. 1991. Foraging and breeding behaviour of the Australian kestrel Falco cenchroides on the northern tablelands of New South Wales. Australian Bird Watcher 14: 85-92.
Paull, D.E. 1981. Buzzard catching stick in flight. British Birds 74: 226-227.
Platt, J.B. 1973. Habitat and time utilization of a pair of nesting sharp-shinned hawks (Accipiter striatus velox) - a telemetry study. M. Sc. Thesis, University of Brigham Young, Provo.
Rabenold, P.P. 1986. Family associations in communally roosting black vultures. Auk 103: 32-41.
Ratcliffe, D. 1980. The peregrine falcon. Berkhamsted; T. & A. D. Poyser:
Real, J. & Mañosa, S. 1986. Independència i dispersió juvenils. In: Real, J. & Mañosa, S. (eds) Pla d'estudi i Protecció de l'Aliga perdiguera Hieraaetus fasciatus (Vicill) a Catalunya. Barcelona; Dep.Biol.Animal (Vertebrats) Un. de Barcelona & C.E.E.M:
Real, J., Mañosa, S., Bertrán, J., Romero, P., Sorolla, A., Fernández, J. & Balaguer, L. 1989. El procés d'independització en els joves d'aliga perdiguera, Hieraaetus fasciatus, al Parc Natural de Sant Llorenç del Munt i l'Obac. In: I Trobada d'Estudiosos de St. Llorenç del Munt i Serra de l'Obac. Barcelona; Diputació de Barcelona:
Romer, M.L.R. 1987. Golden Eagle lifting and dropping stones. British Birds 80: 18.
Rosenfield, R.N. & Kanvik, A. 1983. Precocious nest defense behavior by a sharp-shinned hawk. Raptor Research 17: 62-63.
Schaadt, C.P. & Rymon, L.M. 1982. Innnate fishing behavior by ospreys. Raptor Research 16: 61-62.
Schuyl, V.G., Tinbergen, L. & Tinbergen, N. 1936. Ethologische Beobachtungen am Baumfalken (Falco s. subbuteo L.). Journal für Ornithologie 84: 387-433.
Sherrod, S.K. 1983. Behavior of fledgling peregrines. Ithaca; The Peregrine Fund, Inc:
Simmons, R. 1984. Pre-independence behaviour, morphometrics and trapping of fledgling redbreasted sparrowhawks. Ostrich 55: 158-162.
Snyder, L.L. 1931. Faunal investigation of Long Point and vicinity, Norfolk county, Ontario. Trans. Royal Can. Inst. 18: 117-236.
Spofford, W.R. & Amadon, D. 1993. Live prey to young raptors - incidental or adaptive. Journal of Raptor Research 27: 180-184.
Sunyer, C. 1991. El periodo de emancipación del quebrantahuesos: consideraciones sobre su conservación. In: Heredia, R. & Heredia, B. (eds) El quebrantahuesos (Gypaetus barbatus) en los Pirineos: Características ecológicas y biología de la conservación. Madrid; ICONA: 47-63.
Tachibana, S. & Miura, T. 1988. Observations on juvenile golden eagles during the post-fledging period in the southern Kitakami mountains. Aquila chrysaetos 6: 24-27.
Thorpe, W.H. 1963. Learning and instinct in animals. 2nd Edition. Cambridge; Harvard University Press:
Thorstrom, R.K. 1989. Biología reproductiva, comportamiento y extensión territorial del halcón de monte rayado (Micrastur ruficollis). In: Burnham, W.A., Jenny, J.P.C.W. & Turley (eds) Proyecto Maya: Reporte de Progreso II, El uso de aves de rapiña como indicadores del medio ambiente para diseño y manejo de áreas protegidas y el desenvolvimiento de la capacidad local para la conservación de la naturaleza de América Latina. Boise, Idaho; The Peregrine Fund Inc.: 101-116.
Thouless, C.R., Fanshawel, J.H. & Bertram, B.C.R. 1989. Egyptian vultures Neophron percnopterus and ostrich Struthio camelus eggs - The origins of stone-throwing behaviour. Ibis 131: 9-15.
Tinbergen, L. 1940. Beobachtungen über die Arbeitsteilung des Turmfalken (Falco tinnunculus) während der Fortpflanzungszeit. Ardea 29: 63-98.
Varland, D.E., Klaas, E.E. & Loughin, T.M. 1991. Development of foraging behavior in the American kestrel. Journal of Raptor Research 25: 9-17.
Varland, D.E. & Loughin, T.M. 1992. Social hunting in broods of two and five American kestrels after fledging. Journal of Raptor Research 26: 74-80.
Walker, D.G. 1987. Observations on the post-fledging period of the golden eagle Aquila chrysaetos in England. Ibis 129: 92-96.
Walker, D.G. 1988. The behaviour and movements of a juvenile golden eagle Aquila chrysaetos in England in 1986. Ibis 130: 564-565.
Wallace, M.P. & Temple, S.A. 1983. An evaluation of techniques for releasing hand-reared vultures to the wild. In: Vulture biology and management. Los Angeles; University of California Press:
Wiacek, J. 1989. Obserwacje nad okresem post-pisklecym u Blotniaka stawowego (Circus aeruginosus L.). M. Sc. Thesis, University of Marii Curie-Sklodowskiej, Lublin.
Wyllie, I. 1985. Post-fledging period and dispersal of young sparrowhawks Accipiter nisus. Bird Study 32: 196-198.
Table 1. Hunting behaviour of raptors during the post-fledging dependence period.
Table 1. continued
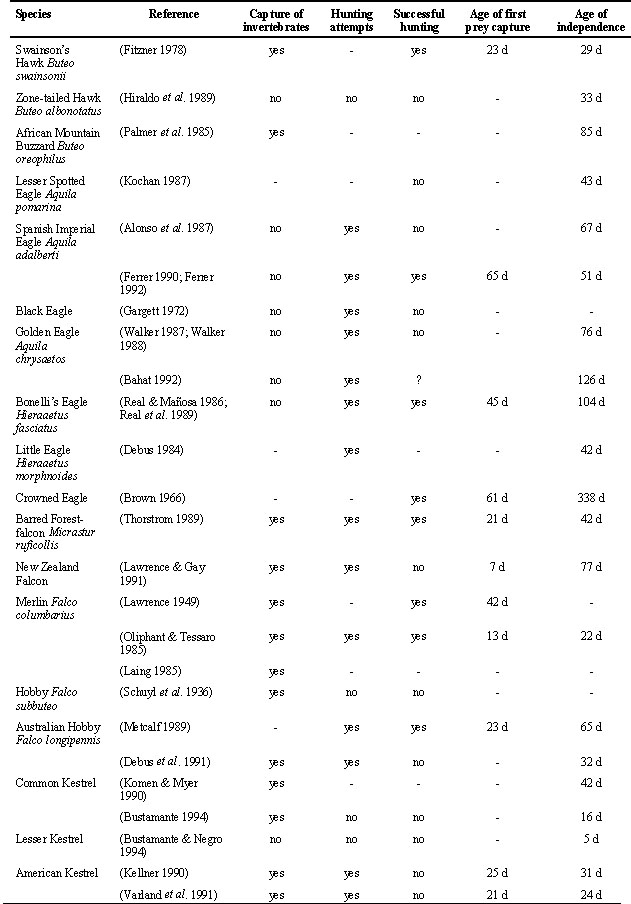
Table 1. continued

Capture of invertebrates: When invertebrates are the first prey captured by fledglings or they have been recorded as prey of fledglings in species in which adults do not normally feed on invertebrates. Hunting attempts: When fledglings try to capture prey without success or are observed chasing habitual prey of the species. Successful hunting: When they have been observed capturing an habitual prey of the species before independence. Age of first prey capture: Mean number of days between first flight and first prey capture. Age of independence: Mean number of days between first flight and independence from parents. (1) They were able to find carrion or dead animals before independence (or capable of breaking bones in the case of the Bearded Vulture), but active hunting was not recorded. (-) No information available on that aspect from the reference. (?) Information is inconclusive on that aspect
Table 2. Behaviour that could be related with adults teaching hunting skills to fledglings.
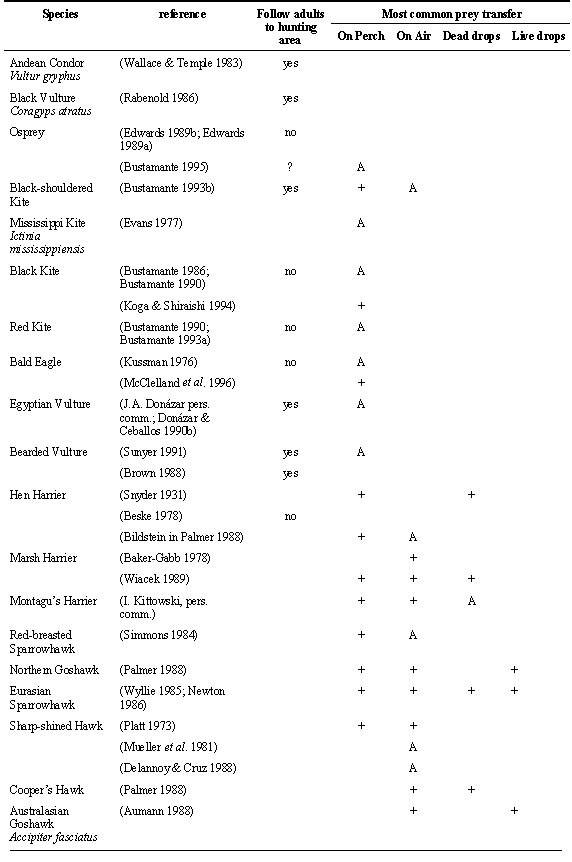
Table 2. continued
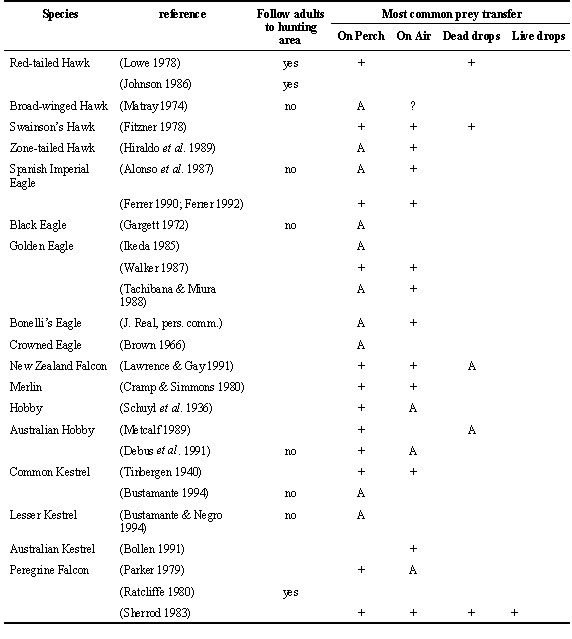
Types of prey transfers: Classified as perch transfer when parent and/or offspring are perched while transfer takes place. Direct prey transfer talon to talon, beak to talon or beak to beak from parent to offspring on the wing are considered air transfers. If air transfers are not clearly described in the source they are classified as this type, that is the most frequent one. ‘Dead-drops’ are transfers in which the parent drops a dead prey in the air that is retrieved by the offspring in the air or on the ground (Sherrod 1983). ‘Live-drops’ are transfers in which the parent drops a live prey that is trapped by the offspring in the air (Sherrod 1983). + = this kind of prey transfer has been recorded. A = All or most prey transfers, once fledglings were able to fly, were of this type. ? = Is not clear if this kind of prey transfer took place.
Table 3. Play behaviour in raptors.
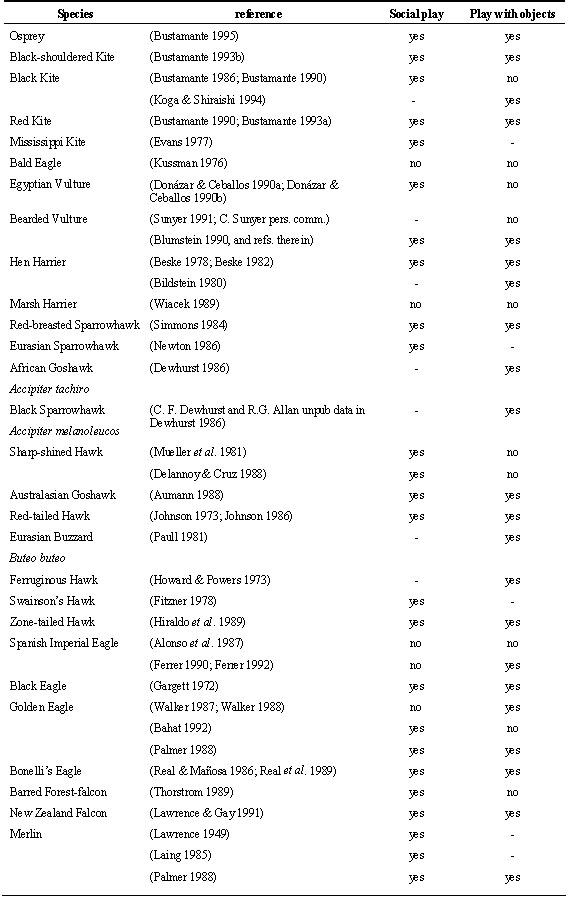
Table 3. continued