S33.2: The migration of land birds over the Pacific Ocean
Timothy C. Williams, Janet M. Williams
Department of Biology, Swarthmore College , Swarthmore, Pennsylvania, 19081 USA.,
fax 1 610 328 8663, e-mail twillia1@swarthmore.edu Williams, T.C. & Williams, J.M. 1999. The migration of land birds over the Pacific Ocean. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1948-1957. Johannesburg: BirdLife South Africa.The migration of land birds over the Pacific Ocean has been studied using distribution and banding data, radar, and ground-based and satellite-based radio telemetry. Migration routes with long over-water components include eastern Pacific movements along the North and South American western coasts, the central Pacific flyway from Alaska and Siberia to Hawaii and the South Pacific islands, and western Pacific flights from the northern Asian coast to the South Pacific and Australia. Non-stop flights appear to approach 8000 km. Radar studies indicate that fixed heading (vector) orientation is sufficient to direct most observed migrations, but satellite tracks and the ability of birds to reliably locate small, remote islands challenges this hypothesis.
INTRODUCTION
The Pacific Ocean is by far the greatest body of water on earth. It stretches 140° in latitude and 150° in longitude at the equator. Yet despite its immense size, land birds cross the Pacific Ocean to a greater extent than any other ocean. Land bird migration is well documented over the North Atlantic Ocean as well as the Mediterranean Sea and many smaller bodies of water, and it is suspected to occur over the northwestern Indian Ocean (Williams & Williams 1990a). Over-water segments of these non-stop flights may be as long as 3500 km. There is no evidence of the migration of land birds over the much larger expanses of the southern Atlantic and Indian Oceans (Williams & Williams 1990a). Flights over the Pacific may be as long as 11000 km of over-water flight with non-stop segments of 6000 km or even 8000 km (Barter & Hou 1990; Marks & Redmond 1994; Piersma & Gill 1998). Birds on such long flights, metabolising at the rate of a human running end to end 4 minute miles for 4 days, represent the most intense energy expenditure known and challenge many of our ideas of flight physiology. Long over-water flights to geographically small goal areas also severely test the known limits of animal orientation. The goals of these migrants are often not large continental land masses but tiny mid-ocean islands many of which are successfully reached by juvenile birds on their first migratory flights unaccompanied by adults.
The great majority of land birds making long over water flights are shorebirds (Charadriiformes) and to a lesser extent waterfowl (Anseriformes), although a significant number of land birds crossing the western North Atlantic are passerines (Passeriformes; Williams and Williams 1990a). For a few species of land birds, long over-water flights may be the primary migration route, but in most cases all indices of migration density indicate that over-land migrations involve many more birds than those over open water. Visual observations at sea and on islands rarely reveal numbers of land birds comparable to those seen on land, and simultaneous radar observations of migration over land and off shore suggest a difference of about two orders of magnitude (Williams et al. 1977; Williams & Williams 1990 a and b, 1994). This does not necessarily indicate that birds regularly fly around the perimeter of bodies of water as large as the western North Atlantic or the western Pacific, only that energetically favourable migration routes over such large bodies of water are rare for land birds (Williams & Williams 1990a).
Studies of the orientation of land birds over the North Atlantic have generally suggested a single migratory route crossing any given body of water (Williams & Williams 1990a). Flights over the Pacific appear to be more complex and take multiple paths, some intersecting at right angles.
METHODS
Due to the remote nature of both the breeding and, in many cases, the non-breeding areas of Pacific migrants, our knowledge of seasonal distribution of migrants in this area is often incomplete especially when compared with the much better studied routes of Palearctic-African or Nearctic-Neotropical migrants. The available data on most Pacific migrant species has been recently summarised by M. Weston in Marchant and Higgens (1993) and Higgens and Davies (1996). We follow their nomenclature in this review. We consider only those species for which there is sufficient data to indicate a migratory route. The exclusion of a species or population from this review should not be taken to indicate a lack of its migratory activity in the Pacific region, only a lack of clear data at the present time.
Land birds are often observed from ships in the Pacific but these observations have rarely been systematic (Casement 1996; Cheshire 1996). Systematic observations from the ships NOAA SURVEYOR and SS UNIVERSE were made as described in Williams and Williams (1994). Observers with 8X or 10X binoculars maintained regular watches on the upper decks of the ship, usually forward. The field of view which varied from 180° to 270° to a radius of 1 km was searched in a regular pattern for birds.
Several different types of radar were used for the observations reported here. Details of the instruments will be found in the publications cited. Observations at Cold Bay, AK, the North American Arctic coast, Anchorage, AK, Guam, Haizhou Bay, and Hong Kong. utilised surveillance radars which detected birds in the range of 20 to 100 km. Observations in Western Australia near 80 Mile Beach were made with a meteorological radar which generally detected birds at less than 50 km range. Altitude information of migrants was obtained from the meteorological radar and from height finding radar at Cold Bay and Guam. The radars used south and east of Anchorage, AK, were modified marine radars with a range of 0.1 to 3 km and were unlikely to detect birds above 500 m.
Two types of radio telemetry data are reported here; see citations for system details. Ground-based telemetry refers to systems in which the receiver is located on the ground or in a light aircraft. The range for these systems is less than 100 km. Satellite-based telemetry uses the ARGOS system to give the location of the transmitter on the earth. This system requires that the transmitter be in line of sight communication with the satellite; position fixes are often infrequent.
Track and groundspeed refer to the velocity of a bird relative to the earth. Heading and airspeed refer to velocity relative to the airmass.
RESULTS
Eastern Pacific
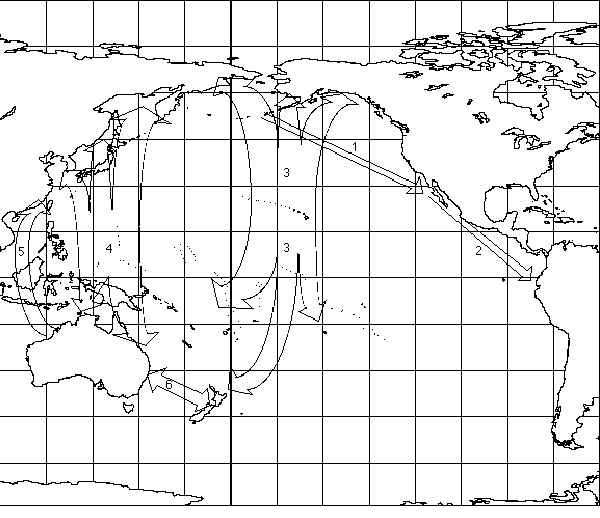
Several species of birds cross the Gulf of Alaska paralleling the North American Coast. Brent Geese Branta bernicla cross more than 4000 km of open water from Izembek Lagoon on the Alaska Peninsula to Baja California (Dau 1992). Based on weather patterns and en route observations, Dau (1992) calculates that the actual flight distance is about 5000 km and takes 60 to 95 hours including any time to rest on the ocean (1 in Fig. 1). Brent Geese tracked for more than 100 km after take off flew at an average altitude of 1100 m (Dau 1992). Cackling Canada Geese Branta canadensis minima appear to make similar offshore flights from the north shore of the Alaskan peninsula to wintering grounds in southern Oregon and central California; banded birds completed the crossing in about 48 hours (Gill 1997).
Morrison (1984) reports several species of shorebirds (Charadriiformes) including Western Sandpiper Calidris mauri and Dunlin C. alpina make long flights off of the North American west coast. Visual observations from the ship NOAA SURVEYOR in the Gulf of Alaska revealed passerines including warblers Parulidae and sparrows Fringillidae moving SE from Alaska toward Oregon (U.S.A.). Observations from the same ship 1000 to 2000 km SW of the North American coast revealed numerous shorebirds, including Golden Plover (probably Pluvialis fulva) and Pectoral Sandpiper Calidris melanotio, moving in a more southerly direction (115° to 220°) toward Hawaii and the southern Pacific islands (Kloeckner et al. 1982; Philip K. Stoddard, Dept. Biological Sciences, Florida International University, Miami FL 33199 USA and Peter Pyle, Point Reyes Bird Observatory, Point Reyes Station, CA 94956 USA pers. comm.). Ground-based radio telemetry of Western Sandpiper revealed that on their spring migration these birds did not make long flights but moved more slowly up the coast using multiple staging areas (Iverson et al. 1996). The pattern of southward non-stop over-water flights and northward coastal flights, common in the western North Atlantic (Williams & Williams 1990a), may also apply to the smaller migrants in the eastern North Pacific.
Wilson's Phalarope Steganopus tricolor appear to fly about 4000 km over water from the southwestern United States or northwestern Mexico to Ecuador or Peru (2 in Fig. 1; Jehl 1988).
Central Pacific
A number of shorebirds including Pacific Golden Plover Pluvialis fulva, Bar-tailed Godwit Limosa lapponica and Bristle-thighed Curlew Numenius tahitiensis, Whimbrel Numenius phaeopus, Wandering Tattler Heteroscelus incanus, Ruddy Turnstone Arenaria interpres, Sanderling Calidris alba, and ducks primarily Northern Pintail Anas acuta and Northern Shoveler A clypeata appear to make flights from staging areas in eastern Siberia and/or western Alaska southward to Hawaii and the southern Pacific islands (Williams & Williams 1990a; Higgens & Davies 1996).
Pacific Golden Plover show remarkable site fidelity in both Hawaiian breeding and Alaskan non-breeding areas (Johnson & Connors 1996). Ground based radio telemetry has been recently used to confirm minimal ground speeds of 50 km/h in flights from Hawaii to Alaska (Johnson et al. 1997). Nineteen radio-tagged birds all landed on the Alaskan Pacific coast well to the east of the great circle route from Hawaii to their breeding grounds. The birds were presumably deflected by westerly winds. Most Pacific Golden Plover over fly the coast continuing to inland breeding areas for a total non-stop flight of at least 5000 km (Johnson et al. 1997).
Williams & Williams (1990a) and Kloeckner et al. (1982) concluded from radar observations that the Hawaiian Islands were a terminus of migration and that birds moving to more southerly non-breeding areas stop and feed at Hawaii. Marks and Redmond (1994) found that 97% of Bristle-thighed Curlew landing on Laysan Island in the northern Hawaiian Islands stay for the season and, thus, concluded that birds moving further south must over fly Hawaii. The nearest islands to Hawaii would require total non-stop flights of 6100 to 7100 km. Further support for very long non-stop flights was obtained by Piersma and Gill (1998) who analyzed the fat content of Bar-tailed Godwits, just before or after take-off on both the northbound and southbound flights between Alaska and New Zealand, and found fat to be ca. 55% of total fresh body mass. The digestive organs of these birds were also greatly reduced.
Indirect evidence suggests that central Pacific migrations are performed at more than 500 m altitude. Radar observations at Anchorage, AK with a radar which detected birds to at least 3000 m altitude revealed light to moderate southward migrations in late August, a time when central Pacific migrants were departing southward. Simultaneous observations with a short range radar (which was unlikely to detect birds above 500 m) on the coast south of Anchorage at Homer and Cordova, AK failed to detect these migrants suggesting that the birds were by then flying at altitudes of more than 500 m, beyond the range of the short range radar (Williams & Williams unpubl.). Systematic visual observations from the SS UNIVERSE in the northern Pacific during both September and May failed to detect any migrant shorebirds although large numbers of seabirds were counted (Williams & Williams 1994). This might be due either to birds flying above the range of visual detection or to the lack of shorebird migrations during the 10 day transit of the area.
Western Pacific
Many shorebird species make regular migrations between eastern Asia or Siberia and Australia or islands of the western Pacific (Williams & Williams 1990a; Marchant & Higgins 1993; Higgens & Davies 1996). For most of these species the route taken is not clear as migrants are seen all along the eastern coast of Asia (Marchant & Higgins 1993; Higgens & Davies 1996). Some of these birds undoubtedly move along the coast but others of the same species make over-water flights of undetermined length. Other species are rarely found on the Asian coast south of Korea, or are regular migrants in the Mariana Islands and, thus, are presumed to make over-water flights from northern Asia to the south Pacific (4 in Fig. 1). These species include the Pacific Golden Plover, Latham's Snipe Gallinago hardwickii, Bar-tailed Godwit, Whimbrel, Grey-tailed Tattler Heteroscelus brevipes, Wandering Tattler, Ruddy Turnstone, Great Knot Calidris tenuirostris, Red Knot C. canutus, Eastern Curlew Numenius madagascariensis, Red-necked Stint C. ruficollis and Sharp-tailed Sandpiper C. acuminata (Barter et al. 1988, Williams & Williams 1990a; Marchant & Higgins 1993; Higgens & Davies 1996; Stinson et al. 1997). Of these the Latham's Snipe, Eastern Curlew, Bar-tailed Godwit, and Great Knot are sufficiently rare in the Marianas to suggest they over fly these islands entirely in a single non-stop flight to or from the south Pacific (Barter et al. 1988; Barter 1989; Barter & Hou 1990; Johnson & Connors 1996; Stinson et al. 1997).
Radar observations at Haizhou Bay near Shanghai revealed relatively small numbers of shorebirds departing the coast to the SSE or SE in autumn. The density of these migrants was about 100 times less than that of other migrants, primarily passerines moving SW along the coast. No significant numbers of birds were seen arriving from the Pacific (from the E or SE) in spring (Mao 1984, 1985; Mao & Zhou 1987; Williams & Williams 1990a). Radar observations from August to November on the island of Guam in the Marianas showed migrants passing over the island to the SSE. The density of birds over flying the island was poorly correlated with increases in birds visually counted on the island suggesting that most migrants over fly the Marianas (Williams & Grout 1985; Williams & Williams 1988). A small number of altitude measurements indicated a maximum altitude of 2500 m (Williams & Williams 1990b). Observations from the SS UNIVERSE on both autumn and spring passages along the Asian coast failed to detect significant numbers of shorebirds again suggesting flight above the limits of visual detection, although the observations in October may have been too late to detect the major trans-Pacific flights (Williams & Williams unpubl.).
Departure of migrants from 80 Mile Beach, Western Australia has been observed both visually and with radar (Lane & Jessup 1985; Minton 1989; Tulip et al. 1994). The great majority of birds were Bar-tailed Godwits and other species whose next staging area on the Asian east coast was north of the departure site. The birds, however, took off consistently to the NW in all years. Tulip et al. (1994) suggest that they may take a more westerly route in spring to take advantage of favourable winds (5 in Fig. 1). Spring radar observations at Hong Kong on the western edge of this route revealed large numbers of birds arriving from the SW to SSE at times of day suggesting 1300 to 2200 km flights over the South China Sea (Myres & Apps 1973; Mellville 1980). Observations in the South China Sea in both autumn and spring from the SS UNIVERSE detected small numbers (11) of unidentified shorebirds, and significant numbers (291) of passerines including 243 Barn Swallows Hirundo rustica predominantly in the spring. It is not clear at present to what extent western Pacific migrants generally utilise an elliptical migration route: over-water southward movements and coastal northward movements as first suggested for Ruddy Turnstone by Thompson (1973).
The Queensland Wader Study Group of Australia and the Wild Bird Society of Japan have successfully satellite-tracked Eastern Curlew from Moreton Bay, Queensland, Australia to their breeding grounds in the Amur River basin in far eastern Russia. Six birds demonstrated their ability to make long, well directed, over-water flights from Moreton Bay over the western Pacific, and one bird has been tracked for an entire migratory cycle, returning to Moreton Bay from the breeding grounds (Musuyuki J. Ueta, Research Center, Wild Bird Society of Japan, 2-35-2 Minamidaira, Hino, Tokyo 191-0041, Japan and Peter Driscoll, Fahey Rd, Mt Glorious, Qld, 4520, Australia pers. comm.).
Other movements
In addition to the routes discussed above, large numbers of migrants from central and eastern Asia reach Australia and New Zealand via the islands of Southeast Asia and Australasia (Marchant & Higgins 1993; Higgens & Davies 1996). As this route is not properly a Pacific migration and as it is difficult to exclude stopovers on these islands this route will not be covered in this review.
One of the more unusual aspects of Pacific migration is that Australia may serve as both breeding and non-breeding grounds. The Double-banded Plover Charadrius bicinctus breeds in New Zealand and shares non-breeding areas with a number of Arctic migrants, but the two migration cycles are reversed with the Double-banded Plover departing to the SE while Arctic migrants are arriving from the N and NW (6 in Fig. 1; Higgins & Davies 1996). Some of these birds regularly winter on remote Norfolk and Lord Howe islands (Endersby 1994). Australian Pratincole Stiltia isabella breed in southeastern Australia and fly north for the non-breeding season (Higgens & Davies 1996).
Even within a single species, subspecies or populations may move in opposite directions at the same time, for example the complex migration patterns within the Pacific Golden Plover (Johnson & Connors 1996). Radar observations in northern breeding grounds suggest extensive east-west movements in late summer which complicate any analysis of the origin of north to south movements (Richardson & Johnson 1981; Flock 1973). Radar observations at Anchorage, AK in August revealed major movements to the S and to the NE often simultaneously. Radar and visual observations by B. Cooper east of Anchorage revealed that the NE movements continue at least 350 km inland indicating a significant NE autumnal migration in south central Alaska. This movement probably then turns southeast to move parallel to the coast. (Brian Cooper, PO Box 249, Forest Grove, OR, 97116 pers. comm.).
DISCUSSION
The Pacific migrations bear many similarities to those in the North Atlantic. The migrants generally fly at great altitudes, make a small number of relatively long flights and show little hesitation at crossing open water (Richardson 1976; Williams & Williams 1990a). The primary differences are the great distances involved, the diversity of routes, and the lack of taxa other than shorebirds and waterfowl in the Pacific except in the South China Sea.
Several lines of evidence now suggest that Pacific migrants regularly make non-stop flights of 5000 to 8000 km (Barter 1989; Barter & Hou 1990; Marks & Redmond1994; Johnson & Connors 1996). In assessing the flight range of birds, a number of physiological models currently in use are inadequate to account for observed flights by shorebirds (Marks & Redmond 1994). The basic assumptions of these models may be incorrect or we may need to look more closely at the effects of wind (Butler et al. 1997). The departure of birds from northern latitudes is usually associated with specific wind and synoptic weather conditions (Thompson 1973; Dau 1992; Marks & Redmond 1994; Butler et al. 1997; Gill et al. 1997). Different species appear to react differentially to similar synoptic conditions. Depending on their destinations, birds depart either just before or just after the passage of a low pressure storm in Alaska (Robert Gill, Jr., U.S. Geological Survey, Alaska Science Center, 1011 East Tudor Rd. Anchorage, AK 99503, pers. comm.). Analysis of migratory flights over the western North Atlantic revealed that wind conditions at departure and en route played a significant role in both the speed and landfall of the migratory flight (Williams et al. 1977; Williams & Williams 1978; Larkin et al. 1979; Stoddard et al. 1983; Williams 1991). In such analyses it is important to use actual measured or geostrophic winds rather than averaged or prevailing winds as birds actively select wind conditions for at least the first portion of their flights. The actual flight path under such wind conditions may depart significantly from a great circle route but may be both energetically favourable and require only fixed heading (vector) orientation (Williams & Williams 1990a). Future investigations in the Pacific might benefit from using flight simulations similar to those used for the Atlantic (Stoddard et al. 1983; Williams 1991). Current work suggests that some species may even exceed 8000 km non-stop flights over open ocean. Preliminary analyses by R. Gill, C. Pennycuick, and T. Piersma indicate that the extremely high fat loads and reduction of digestive and reproductive organs of Bar-tailed Godwits, when coupled with favourable in-flight wind conditions, could allow these birds to make an 11000 km non-stop flight from Alaska to New Zealand (R. Gill ibid, pers. comm.).
The distances covered and the increasing knowledge of the site specificity of non-breeding areas severely challenges our understanding of orientation mechanisms involved. Williams and Williams (1990a) were able to assert that Pacific migrations as well as those in the Atlantic could be explained by fixed heading (vector) orientation. Birds appeared to maintain heading under different wind conditions without compensation for wind drift. Goal areas were all relatively large and although individual islands might be small they were grouped into chains, such as the Hawaiian Islands, more than 1000 km long. These conclusions still apply to the great majority of Pacific migrants, especially those moving between the central Asian coast and Australasia. Bristle-thighed Curlew maintaining a suitable fixed heading might be reasonably expected to intersect the Hawaiian Islands which subtend an angle of 45° (on great circle routes) from their staging area in western Alaska. They could then search among those islands, as Kloeckner et al. (1982) estimated Pacific islands could be detected by shorebirds at a distance of 300 km. The same species over flying the Hawaiian Islands to the northern Line Islands would be aiming at a target which subtends only 8° from western Alaska. It seems unlikely that fixed heading orientation would be sufficient to successfully guide a flight to such a small goal area, and it does not seem likely that the birds could search extensively without feeding after a 7000 km flight. Satellite radio telemetry will be important in resolving these issues. Of particular importance will be the behaviour of birds deflected from their flight path by storms. Johnson et al. (1997) and Gill et al. (1997) found that birds could compensate for such deflection after making landfall, but satellite tracking should be able to indicate whether migrants can make such corrections en route. Homing experiments and satellite radio telemetry of Wandering Albatross Diomedea exulans and Laysan Albatross D. immutabilis indicate that these birds may be capable of true bicoordinate navigation (Fisher 1971; Kenyon & Rice 1958; Jouventin & Weimerskirch 1990; Weimerskirch et al. 1993; Nicholl et al. 1995); perhaps similar results will be found for long distance Pacific migrants. Even if we do learn that migrant shorebirds have the capacity for bicoordinate navigation, explaining the orientation of first year birds will present even greater challenges. First year birds appear to migrate apart from the adults (Marchant & Higgins 1993; Marks & Redmond 1994; Higgins & Davies 1996; Johnson & Connors 1996). We know of no mechanism which would allow first year birds to know the coordinates of their non-breeding area even if they were capable of bicoordinate navigation.
The wide and complex dispersal patterns of many species within the Pacific area also represent a challenge to evolutionary biology. In contrast to the Nearctic and the western Palearctic, the breeding areas of Pacific migrants in eastern Siberia and western Alaska remained relatively ice free during the last glaciation (Williams & Webb 1996). These breeding areas were probably more constant in location than breeding areas subject to glaciation which must have shifted extensively with the ice caps. The non-breeding areas, especially those on small Pacific islands, must have changed greatly with the rise and fall of sea level by 100 m or more. The relatively small breeding areas, the wide dispersal for the non-breeding season, and the high vagrancy rates presently observed for many species may be an adaptation to these conditions. Philopatric behaviour on the breeding grounds would be adaptive. Philopatry to non-breeding areas as observed by Marks and Redmond (1994) and Johnson and Connors (1996) deserves more investigation. As an example, the Wandering Tattler is a monotypic species, breeds in far eastern Siberia and Pacific coastal Alaska and in the non-breeding season may be found over much of the Pacific rim from the Americas to New Zealand and Australia, Hawaii and South Pacific islands (Higgins & Davies 1996). It would be of great interest to know whether there are in fact subpopulations within the species with specific migration routes to the widely distributed non-breeding areas or whether, as the name implies, individual birds lack such specific goal areas. We also need to develop explanations of long non-stop flights, especially those that pass over suitable
stopover sites. Piersma (1989) has shown that it is always energetically favourable to make several short flights rather than one long one so other factors such as predation risk must be considered.
Although additional research is necessary to resolve these important questions it must be combined with major efforts at preservation of Asian and Pacific avian habitats, especially those of shorebirds. Many populations of many species covered in this report are threatened, especially in critical stop-over habitats on Asian and Pacific islands (Marchant & Higgins 1993; Higgins & Davies 1996; Gill et al. 1994; Wells & Mundkur 1996).
ACKNOWLEDGEMENTS
We thank the following people for assistance in this review: M. Barter, B. Cooper, C. Dau, P. Driscoll, I. Endersby, R. Gill Jr., O. Johnson, J. Jonkel, H. McClure, T. Mundkur, M. Ueta, P. Walbridge, N. Warnock, M, Weston, G. Wiles; M. Spencer provided bibliographic assistance.
REFERENCES
Barter, M. 1989. Bar-tailed Godwit Limosa lapponica in Australia. Part 1: Races, breeding areas and migration routes. Stilt 14: 42-48.
Barter, M. & Hou, W.T. 1990. Can waders fly nonstop from Australia to China? Stilt 17: 36-37.
Barter, M., Jessop, A. & Minton, C. 1988. Red Knot (Calidris canutus rogersi) in Australia. Part 1: Sub-species confirmation, distribution and migration. Stilt 12: 29-32.
Butler, R.W., Williams, T.D., Warnock, N. & Bishop, M.A. 1997. Wind assistance: A requirement for migration of shorebirds? Auk 114: 456-466.
Casement, M. 1996. 50 years of landbird observations as sea. Sea Swallow 45: 20-23.
Cheshire, N.G. 1996. RNBWS in the Pacific. Sea Swallow 45: 26-32.
Dau, C. 1992. The fall migration of Pacific Flyway Brent Branta bernicla in relation to climatic conditions. Wildfowl 43: 80-95.
Endersby, I. 1994. Vagrancy, or another migration path, of the Double-banded Plover Charadrius bicinctus? Stilt 24: 20-23.
Fisher, H.I. 1971. Experiments on homing in Laysan Albatrosses Diomedea immutabilis. Condor 73: 389-400.
Flock, W.L. 1973. Radar observations of bird movements along the arctic coast of Alaska. Wilson Bulletin. 85: 259-275.
Gill, R.E. Jr., Butler, R.W., Tomkovich, P.S., Mundkur, T. & Handel C.M. 1994. Conservation of North Pacific shorebirds. Transactions of the North American Wildlife and Natural Resources Conferences 59: 63-78.
Gill, R.E., Babcock, C.A., Handel, C.M., Butler, W.R. Jr. & Raveling, D.G. 1997. Migration, fidelity, and use of autumn staging grounds in Alaska by Cackling Canada Geese Branta canadensis minima. Wildfowl 47: 42-61.
Higgens, P.J. & Davies, S.J.J. (eds). 1996. Handbook of Australian, New Zealand and Antarctic Birds Vol. 3: Snipe to Pigeons. Melbourn, Oxford University Press: 1028pp.
Iverson, G.C., Warnock, S.E., Butler, R.W., Bishop, M.A. & Warnock, N. 1996. Spring migration of Western Sandpipers along the coast of North America: A telemetry study. Condor 98: 10-21.
Jehl, J.R. Jr. 1988. Biology of the Eared Grebe and Wilson's Phalarope. Studies in Avian Biology. 12: 1-74.
Johnson, O.W. & Connors, P.G. 1996. Pacific Golden-Plover (Pluvialis fulva). In: Pool, A. & Gill, F. (eds) The Birds of North America No. 201-202; Phildelphia; The Academy of Natural Science, and Washington, D.C.; The American Ornithologists' Union: 1-40.
Johnson, O.W., Warnock, N., Bishop, M.A., Bennett, A.J., Johnson, P. & Kleinholz, R.J. 1997. Migration by radio-tagged Pacific Golden-Plovers from Hawaii to Alaska, and their subsequent survival. Auk 114: 521-524.
Jouventin, P. & Weimerskirch, H. 1990. Satellite tracking of Wandering Albatrosses. Nature 343: 746-748.
Kenyon, K.W. & Rice, D.W. 1958. Homing of the Laysan Albatrosses. Condor 60: 3-6.
Kloeckner, P.D., Williams, J.M. & Williams, T.C. 1982. Radar and visual observations of transpacific migrants. ´Elepaio 42: 77-80.
Lane, B. & Jessop, A. 1985. Tracking of migrating waders in North-western Australia using meteorological radar. Stilt 6: 17-28.
Larkin, R.P., Griffin, D.R., Torre-Bueno, J.R. & Teal, J. 1979. Radar observations of bird migration over the western North Atlantic Ocean. Behavioral Ecology and Sociobiology 4: 225-264.
Mao, Y. 1984. Observation of bird migration by radar in Haizhou Bay, China. M.S. Thesis. Shanghai, East China Normal University.
Mao, Y. 1985. Wader migration in Haizhou Bay, eastern China. Interwader Newsletter 6: 10-11.
Mao, Y. & Zhou, B. 1987. [Observations on Bird migration by radar in Haizhou Bay] Chinese, Eng. Summary. Acta Zoologica Sinica 33: 277-284.
Marchant, S. & Higgins, P.J. (eds) 1993. Handbook of Australian, New Zealand and Antarctic Birds Vol. 2: Raptors to Lapwings. Melbourn, Oxford University Press: 984pp.
Marks, J.S. & Redmond, R.L. 1994. Migration of Bristle-thighed Curlews on Laysan Island: Timing behavior and estimated flight range. Condor 96: 316-330.
Melville, D. 1980. Bird migration through Hong Kong observed by radar and its importance for birdstrike control. Report to Birdstrike Research Unit, Agriculture and Fisheries Departmentt., Hong Kong.
Minton, C. 1989. N.W. Australia wader expedition March/ April 1988. Stilt 14: 6-13.
Morrison, R.I.G. 1984. Migration systems of some new world shorebirds. In: Burger, J. & Olla, B. L. (eds) Shorebirds, migration and foraging behavior. Vol. 6; Behavior of marine animals, New York; Plenum:125-202.
Myres, M.T. & Apps, R.F. 1973. Migration of birds over the south coast of China recorded by radar. Nature 241: 552.
Nicholl, D., Murray, D., Battam, H., Robertson, G., Moors, P., Butcher, E. & Hildebrandt, M. 1995. Satellite tracking of the Wandering Albatross Diomedea exulans around Australia and in the Indian Ocean. Emu 95: 223-230.
Piersma, T. 1989. Hop, skip or jump? Constraints on migration of arctic waders by feeding, fattening and flight speed. Stilt 14: 38-40.
Piersma, T. & Gill, R. Jr. 1998. Guts don't fly: Digestive organs in obese Bar-tailed Godwits. Auk 115: 196-203.
Richardson, W.J. 1976. Autumn migration over Puerto Rico and the western Atlantic: a radar study. Ibis 118: 309-331.
Richardson, W.J. & Johnson, S.R. 1981. Waterbird migration near the Yukon and Alaska coast of the Beaufort Sea: I: Timing, routes and numbers in spring. Arctic 34: 108-121.
Stinson, D.W., Wiles, G.J. & Reichell, J.D. 1997. Occurrence of migrant shorebirds in the Mariana Islands. Journal of Field Ornithology 68: 42-55.
Stoddard, P.K., Marsden, J.E. & Williams, T.C. 1983. Computer simulation of autumnal bird migration over the western North Atlantic. Animal Behavior 31: 173-180.
Thompson, M. 1973. Migratory patterns of Ruddy Turnstones in the central Pacific region. Living Bird 12: 5-23.
Tulip, I., McChesney, S. & De Goeij, P. 1994. Migratory departures from north-western Australia: Behavior timing and possible migration routes. Ardea 82: 201-221.
Weimerskirch, H., Samolard, M., Sarrazin, F. & Jouventin, P. 1993. Foraging strategy of Wandering Albatross through the breeding season; A study using satellite telemetry. Auk 110: 325-342.
Wells, D.R. & Mundkur, T. (eds). 1996. Conservation of migratory waterbirds and their wetland habitats in the East Asian Australasian Flyway. Proceedings of an international workshop, Kushiro, Japan. 28 Nov- 3 Dec 1994. Kuala Lumpur and Tokyo. Wetlands International-Asia Pacific, Publication No. 116 and International Waterfowl And Wetlands Bureau - Japan Committee: 104pp.
Williams, J.M. & Williams, T.C. 1994. Seabirds observed on two global circumnavigations in tropical and subtropical oceans and the subarctic Pacific. Bird Populations 2: 1-22.
Williams, J.M. & Grout, P.C. 1985. Migrants on Guam, Fall l983. ´Elepaio 46: 41-44.
Williams, T.C. 1991. Constant compass orientation for North American autumnal migrants. Journal of Field Ornithology 62: 218-225.
Williams, T.C. & Webb, T. III. 1996. Neotropical bird migration during the ice ages: orientation and ecology. Auk 113: 105-118.
Williams, T.C. & Williams, J.M. 1978. Orientation of transatlantic migrants. In: Schmidt-Koenig, K. & Keeton, W. (eds) Proceedings pf a Symposium onAnimal Migration, Navigation, and Homing. Proceedings in the Life Sciences, Berlin; Springer-Verlag: 239-251.
Williams, T.C. & Williams, J.M. 1988. Radar and visual observations of autumnal (southward) shorebird migration on Guam. Auk 105: 460-466.
Williams, T.C. & Williams, J.M. 1990a. The orientation of transoceanic migrants. In: Gwinner, E., (ed.) Bird migration: physiology and ecophysiology. Berlin; Springer Verlag: 9-21.
Williams, T.C. & Williams, J.M. 1990b. Open ocean bird migration. IEE Proceedings 137, Pt F: 133-137.
Williams, T.C., Williams, J.M., Ireland, L.C. & Teal, J.M. 1977. Autumnal bird migration over the western North Atlantic Ocean. American Birds 31: 251-267.
Fig.1. Principal Pacific migration routes involving long non-stop over-water flights. Arrows indicate hypothetical migration routes; numbers refer to text. Latitude and longitude grid in 15° increments.