S50.4: Holocene bird subfossils from the sites of Ampasambazimba, Antsirabe and Ampoza, Madagascar: Changes in the avifauna of south central Madagascar over the past few millennia
Steven M. Goodman
Department of Zoology, Field Museum of Natural History, Roosevelt Road at Lake Shore Drive, Chicago, Illinois 60605, USA, e-mail goodman@.fmnh.org
Goodman, S.M. 1999. Holocene bird subfossils from the sites of Ampasambazimba, Antsirabe and Ampoza, Madagascar:Changes in the avifauna of south central Madagascar over the past few millennia. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 3071-3083. Johannesburg: BirdLife South Africa.Information is reviewed on bird subfossils excavated at three sites on Madagascar. On the basis of radiocarbon analysis this material dates from the end of the Late Pleistocene to the Recent era. Two of the sites studied, Ampasambazimba and Antsirabe, are on the Central High Plateau, and the third site, Ampoza, is along the southwestern margin of the Central High Plateau. A significant proportion of the bird bones identified from these sites are extinct (33%). When extinct species and extant species that have locally disappeared from each region are combined, 50-80% of the birds recorded as subfossils no longer occur in the immediate vicinity of these sites. Various factors that might account for this high species turnover in a relatively short period of time are discussed.
INTRODUCTION
Several palynological studies published within the past decade have presented evidence of considerable climatic changes on Madagascar during the Holocene (see Burney 1997 for a review). There is increasing evidence that these climatic shifts are strongly implicated in the extinction of numerous vertebrate species (Burney et al. 1997), and in a temporal sense the human factor often cited for the megafauna extinction may have not become a critical factor until after populations of these animals were already on the decline. Current data suggests that, at least in the southwestern portion of the island, these climatic shifts involved a mesic period starting about 5000 yr BP and an arid period between 3500 and 2500 yr BP (Burney 1993). These proposed shifts are mirrored in changes of species representation and habitat types of bird subfossils excavated from sites in southwestern Madagascar (Goodman & Rakotozafy 1997). Several bird species may have gone extinct during this period, rather than after the initial human colonization of the island about 2,000 years ago (MacPhee & Burney 1991; Dewar & Wright 1993). In light of modern conservation problems on Madagascar, one of the important aspects of paleontological studies of Holocene material is that it may allow us to tease apart the factors resulting in faunistic changes and to assess what role natural climatic shifts may have played as compared to anthropogenic perturbations.
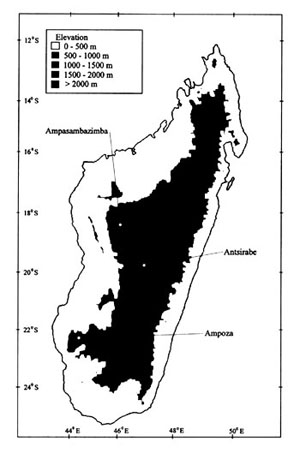
Information on the bird subfossils excavated from three sites, including two on the Central High Plateau (Ampasambazimba and Antsirabe) and one in the southwest at the edge of the Central High Plateau (Ampoza), are presented in this paper (Fig. 1). Most of this material was unearthed before the advent of controlled paleontological techniques, and thus lacks stratigraphic context. This limits interpretations of faunal associations, horizons of deposition and taphonomy. Radiocarbon dates are available of bone material recovered from these sites (Table 1), as is some information on the bone-bearing beds. The modern regions of Madagascar where these sites are found currently contain few large areas of natural habitat, particularly fresh water lakes and large blocks of forest. The regions of the high plateau surrounding the sites of Ampasambazimba and Antsirabe are composed of vast areas of anthropogenic grasslands and wetlands converted to rice paddy. Some natural habitat still remains in the Ampoza region, which forms a transitional zone between humid and deciduous forests. The subfossil material provides insight to the former habitat and biota that occurred in these areas.
METHODS
Collections
The material used in this study is housed in four museums: AMNH - American Museum of Natural History, New York; BMNH - The Natural History Museum, Paleontology Section, London (formerly British Museum [Natural History]); MNHN - Laboratoire de Paléontologie and Laboratoire d'Anatomie Comparée, Muséum National d'Histoire Naturelle, Paris; and UA - Service de Paléontologie, Université d'Antananarivo (to where a large number of bird subfossils from the Académie Malgache was transferred).
Sites
Ampasambazimba
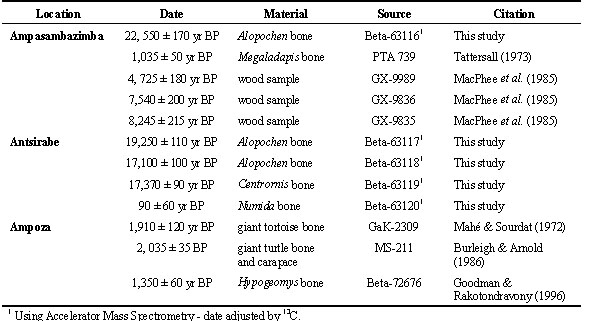
Ampasambazimba, located on the Central High Plateau at 18°39'S, 46°01'E, is one of the best known subfossil sites on the island and excavations conducted sporadically between 1902 and 1984 (Standing 1908; Lamberton 1934; Tattersall 1973; MacPhee et al. 1985) have unearthed an enormous amount of bone material. These include, for example, 15 species of lemur of which seven are extinct (Godfrey et al. 1997). The bird bone collections from this site are housed in the MNHN and UA. Some of the material in the MNHN that came from Ampasambazimba and near Antsirabe arrived at the museum in the same shipment and were accessioned together; a significant portion of these bones lack any information on provenance and can only be recognized as coming from Ampasambazimba or the Antsirabe region (including Betafo). The species represented in this mixed material are not presented in this paper. The majority of the bone bearing horizons excavated during the early excavations at Ampasambazimba were below a 1 m level of fine black humus, inferior to which was a layer of forest debris, followed by the bone bearing beds mixed with plant debris and wood (Standing 1908). Radiocarbon dates from the site include a small sample of bone fragments of the extinct shelgoose Alopochen sirabensis (UA 360) determined by Accelerator Mass Spectrometry (AMS) at 22,550 ± 170 yr BP, and a portion of a tibia diaphysis (BM M 9927) of an extinct giant lemur Megaladapis grandidieri that yielded a date of 1035 ± 50 yr BP (Tattersall 1973).
Antsirabe
The subfossil locality ‘Antsirabe’ is composed of several sites around the Antsirabe and Betafo area (Fig. 1) on the central portion of the high plateau. Jully (1898) reviewed information on pre-1900 excavations made near Antsirabe. Bird material in the MNHN and UA include specimens collected by Bastard, Bouet, A. and G. Grandidier, Hildebrandt, Müller, and Rosaas, as well as material sent to these museums by the Académie Malgache. The stratigraphic sequence of deposits excavated in 1898 near Andrangy, west of Antsirabe, consisted of (from the surface) rich black soil deposits, a layer of peat (Tourbe consommée), peat containing non-decomposed vegetation and bone-bearing deposits, red soils with vegetative matter, and finally a layer of gravel (Jully 1898). This is in contrast to a site in the Vallée des Eaux, excavated during the same season and a few km from Andrangy, with the following sequence (from surface): a layer of decomposing vegetation, peat with vegetative matter, a layer of wet peat, gravel with the bone bearing deposits, and a calcareous deposit.
The BMNH holds the collection made by Forsyth Major at ‘Sirabé.’ Andrews (1897) previously reported on most of the bird material from this collection and described several extinct species. The bones excavated by Forsyth Major were obtained from numerous sites, often from a depth of 3.6 to 4.5 m. The stratigraphic sequence appears to have been a surface layer, 1.5 to 1.8 m deep, of rich black earth, generally containing little bird bone, followed by coarse gravel and calcareous conglomerate with broken bone. The principal fossil-bearing layer was found below the conglomerate in a marly deposit (Andrews 1897). Several bone samples from the site have yielded Radiocarbon dates using AMS techniques: Alopochen sirabensis (UA 142) - 19,250 ± 110 yr BP; A. sirabensis (UA 119) - 17,100 ± 90 yr BP; the extinct shelgoose Centrornis majori (UA 214) - 17,370 ± 90 yr BP; and Helmeted Guineafowl Numida meleagris (UA 322) - 90 ± 60 yr BP (Table 1).
Ampoza
In 1929 E. I. White, a paleontologist from the British Museum (Natural History) and associated with the Franco-Anglo-Américan zoological expedition to Madagascar, excavated the site of Ampoza (Fig. 1), about 27 km east of Ankazoabo (White 1930). The majority of the Ampoza material was deposited in the MNHN. White's Ampoza excavation was conducted near subfossil sites worked by Lamberton (1934) and Ljungqvist (Wiman 1935). Some information is available from White's excavation sites (White 1930): ‘The first locality was in the valley of a small stream....Here a trial pit had already been dug, and fortunately the overburden, consisting of about 8 feet [=2.4 m] of black earth, had been removed from a considerable area. However, there was then an 18-inch [46 cm] layer of limestone to be cleared away over most of it, and this presented much difficulty at times. Underneath the limestone was the fossiliferous layers consisting of two or three feet of black mud and finally barren light-coloured sands, sometimes coloured bright blue and green.’ In the middle of a second pit he excavated, which was about 40 m long and 20 m wide, was a fresh water stream that was approximately 30 m from the Ampoza River.
Radiocarbon dates are available from the site that include an AMS date from a tibia (MNHN 1931.6) of Hypogeomys antimena, an endemic large rodent that no longer occurs in the region, which yielded a date of 1350 ± yr BP (Goodman & Rakotondravony 1996). Two dates are available for bone remains of giant extinct tortoises from the site: 1,910 ± 120 yr BP (Mahé & Sordat 1972) and 2,035 ± 35 yr BP (Burleigh & Arnold 1986; Table 1).
RESULTS
Thirty-five different bird species were identified from bone remains recovered from the three subfossil sites: 13 from Ampasambazimba, including eight extinct species; 16 from Antsirabe, including six extinct species; and 20 from Ampoza, including five extinct species (Table 2). Twelve species of extinct birds were found at these sites, 34% of the total represented avifauna. This figure is conservative since several species of elephant birds (Aepyornis and Mullerornis) probably are represented at these sites (Lamberton 1934; Wiman 1935); however, due to difficulties in defining the species' limits of members of these genera no specific determinations have been presented herein.
Twenty-three of the species present at these sites use or used aquatic ecosystems as their main habitat (Table 2). For the extinct forms, the assignment of presumed habitat type relies on extrapolation with extant congeners or inference of functional morphology based on osteology. The habitat used by the extinct lapwing Vanellus madagascariensis is not clear since members of this genus are both aquatic and terrestrial (Johnsgard 1981); however, it has been placed in the aquatic category. Of these 23 aquatic species, five are extinct. Further, for the majority of extant species, their modern distribution no longer includes the Central High Plateau or the general Ampoza area (Table 2).
DISCUSSION
General Considerations
The taxa identified from bone remains recovered from the various subfossil sites clearly represent a portion of the species that formerly lived in each region. Thus, little can be gleaned from direct faunistic comparisons of the species composition at each site. Perhaps a more fruitful analysis involves habitat utilization of the represented taxa, percentages of animals extinct, and changes in distribution of extant species.
Eight of the 13 species identified from Ampasambazimba are extinct, as compared to six of 16 at Antsirabe, and five of 20 at Ampoza (Table 3). At Ampasambazimba slightly more than half of these species are terrestrial and at the other two sites one-half to two-thirds are aquatic. There are substantial differences between the sites in the habitat preferences of extinct species: at Ampasambazimba terrestrial species dominate, at Antsirabe aquatic species are more common, and at Ampoza aquatic and terrestrial species are nearly equal in representation. The greatest shift in the modern distribution of species found at these sites but no longer occurring in the region is at Ampoza, where over half of the identified extant species identified in the remains currently are unknown from this area of the island. The majority of these species are found in the western or northwestern portions of Madagascar. When island wide and regional extinctions are combined, between 50 and 80% of the species identified at the subfossil sites no longer occur in the surrounding region. Thus, the degree of change of the island's avifauna in a relatively short period is rather dramatic. Further, when figures of extinct non-avian taxa recovered from each of these sites are included (Godfrey et al. 1997), it is abundantly clear that there has been massive change in the vertebrate communities of Madagascar since the end of the late Pleistocene.
A considerable debate has been waging in the literature about the origins of these extinctions and cause of geographical range contractions of the endemic organisms of Madagascar. The explanation for the high extinction rate on the island since the beginning of the Holocene probably lies somewhere between the extremes of natural environmental change and anthropogenic modifications of the environment (see review by Dewar 1997). Given the lack of stratigraphic control associated with the excavated bird bone material at the three sites discussed in this paper, it is impossible to reconstruct the sequential shifts in the biota through time and to correlate these changes with climatic or anthropogenic factors.
During the early excavations at Ampasambazimba, evidence was found to potentially implicate humans in the deposition of a portion of these bone deposits. This evidence includes a modified elephant bird tibiotarsus, a wood tool, and an ceramic pot (Fontoyont 1909); however, this extrapolation is weak and not acceptable (Barthère 1915; MacPhee et al. 1985). More recent excavations at the site found no evidence of human presence or context in the subfossil-bearing strata (MacPhee et al. 1985). At Antsirabe, Jully (1898) reported finding a pottery fragment 0.2 m below bone-bearing deposits. In the absence of a detailed stratigraphic context, it is impossible to implicate humans in the deposition of the large quantities of bone material recovered from these sites.
The temporal breadth of radiocarbon dates available from the material associated with subfossil deposits recovered at Ampasambazimba and Antsirabe ranges from Late Pleistocene to the Recent era (Table 1). For example, the date of 1035 ± 50 yr BP presented by Tattersall (1973) for bone material of the giant lemur Megaladapis shows that this animal lived contemporaneously with humans on the island. Further, the recent date of 90 ± 60 yr BP for a guineafowl bone (Numida) from Antsirabe clearly demonstrates that modern material is included within these deposits. Guineafowl may have been introduced to Madagascar from east Africa (Langrand 1995). Until these sites are reopened and human artifacts are recovered in a controlled stratigraphic context, it is difficult to construct any association between humans, the deposition of the bones of animals, and the causality of their extinction.
The case of Alopochen
Bones of Alopochen sirabensis are some of the most common in the bird material excavated at Ampasambazimba, Antsirabe, and Ampoza. All of the c-14 dates available from the first two sites range from approximately 17,000-22,500 yr BP (Table 1). Material of this species from Beloha in the extreme southwest yielded a radiocarbon date of 1380 ± 90 yr BP (Goodman & Rakotozafy 1997), thus confirming this species continued existence on Madagascar until after human colonization of the island. The only extant member of this genus is the Egyptian Goose A. aegyptiacus, which is widespread on the African continent (Brown et al. 1982).
Currently, the most common species of large waterfowl on the island is the Knob-billed Duck Sarkidiornis melanotos, which occurs rather commonly in the small pot-hole lakes in the Ampoza region. Bones of Sarkidiornis are rare at these subfossil sites. Several characters clearly differentiated the long bones of Sarkidiornis and Alopochen (Rakotozafy 1993). On the African continent the distributions of S. melanotos and A. aegyptiacus overlap considerably, and there is no clear evidence of competition between these species (Brown et al. 1982; Lewis & Pomeroy 1989). Given these factors it would appear that either Sarkidiornis has only recently colonised Madagascar or did not occur in the same habitat as A. sirabensis. The island of Mauritius also was occupied by another extinct species of shelgoose, A. mauritianus, which apparently existed into historical times (Cheke 1987). Alopochen aegyptiacus seems able to withstand relatively heavy hunting pressure and human persecution. The reasons for the extinction of A. sirabensis and A. mauritianus are not clear.
The case of Anas bernieri
Bernier's Teal Anas bernieri is a duck endemic to Madagascar that now is classified as vulnerable to extinction (Langrand & Wilmé 1993). The total current population has been estimated as probably not exceeding 500 individuals (Safford 1993). The modern distribution of this species appears to be limited from the central western coast near Morombe north to the Ambanja region. On the basis of bones identified as A. bernieri recovered from Ampasambazimba, Antsirabe, Ampoza, and numerous sites in the extreme southwestern portion of the island (Goodman & Rakotozafy 1997), it is clear that this species until relatively recently had a much broader distribution across the island.
GENERAL CONCLUSIONS
In a review of recent extinctions on Madagascar, Dewar (1997) emphasised that global explanations for the events that resulted in the massive environmental changes on the island over the Holocene are difficult to construct and that each region, with its different cultural and biological factors, needs to be considered separately. Thus, the factors that lead to the extinctions and shifts in the bird communities near Ampoza and the Central High Plateau sites of Ampasambazimba and Antsirabe may not have been the same.
There is evidence that over the past few millennia southwestern Madagascar has become distinctly drier. The dramatic results of this aridification are supported by the extinction or local disappearance of numerous animals that require mesic environments (MacPhee 1986; Stuenes 1989; Goodman & Rakotozafy 1997). Burney (1993) analysed a pollen core, representing a 5,000 year environmental sequence, from a coastal area north of Toliara, 140 km SW of Ampoza. His results indicate that until about 3,000 years ago the southwestern portion of the island was distinctly wetter, and during the following millennium there is good evidence of aridification. At approximately 1900 yr BP there was a substantial increase in grass pollen and a correlated decrease in woody plant pollen. Thus, in the southwest, just before human colonization of the island, there was significant habitat modification. Burney's scenario of ecological change is supported by the composition and species' habitat requirements of bird subfossils recovered from a variety of sites in the extreme southwest. Ampoza is not too far from these other sites and probably experienced the same natural ecological change. However, on the basis of radiocarbon dating a giant terrestrial tortoise, which is now extinct, still occurred near Ampoza during this hypothesised period of ecological change, and Hypogeomys, a large terrestrial rodent that currently has a very restricted geographical range in the Morondava area, continued to exist for at least several hundred years more (Table 1). Little in the way of large freshwater ecosystems remain today in the general Ampoza area, and given that 75% of the species identified from the site are aquatic and that 55% of them are still extant but no longer occurring in the region would support a model of ecological change. Significant human migration into the interior portions of the southwest is thought to have occurred several hundred years after initial colonisation of the island, and dramatic anthropogenic ecological change in the region may not have commenced until considerably later (Dewar & Wright 1993). In the general Ampoza area there has been little conversion of the remaining aquatic ecosystems into rice paddy.
The explanation for the disappearance of numerous species of animals on the Central High Plateau is not as clear as in the case of Ampoza. Pollen data and radiocarbon dating based on a core taken in Lake Kavitaha, not far from Ampasambazimba, indicate that starting in the oldest portion of the core (1490 ± 60 yr BP) there was a distinct reduction in woody pollen and a proportional increase in grass pollen (Burney 1987a). Whether these shifts are associated with natural environmental change or human modifications of the local environment is open to question. However, soon thereafter, approximately 1400 yr BP, the amount of charcoal in the samples increased substantially, and this probably signals the start of anthropogenic environmental change. Supportive radiocarbon dates associated with archaeological sites on the Central High Plateau indicate that the first signs of human modification of the region date from the 7th-century, settlements probably were established in the 12th-century, and that there was a significant augmentation in anthropogenic modifications of the environment for agricultural or pastoral purposes in the 18th-century (Wright et al. 1992). As far as aquatic ecosystems are concerned, the major habitat for waterbirds, the principal anthropogenic influence probably would have been conversion of marshes and lake ecosystems into rice fields. The first evidence of the cultivation of rice on the Central High Plateau is from the 15th-century (Wetterstrom & Wright 1992). Certainly hunting pressure on waterbirds and the raiding of nests for eggs would have commenced at an earlier date. There appears to be no textual evidence from early European visitors to Madagascar that large and notable waterfowl, such as the enormous Centrornis majori, still were in existence on the island during the 16th and 17th centuries (e.g. Flacourt 1658). However, this negative evidence cannot be used to say that these birds already were extinct at the time.
The earliest portion of the Lake Kavitaha core was dated to about 1500 yr BP, and thus is not useful to address questions about local environmental change in the Ampasambazimba region before human colonization of Madagascar. Other pollen cores taken in Lake Tritrivakely, at 1800 m in the hills above Antsirabe, provide a record of the local environmental conditions over the past 36000 years (Burney 1987b; Gasse et al. 1994). Perhaps due to the nature of the type of vegetation occurring in high mountain zones, traces of clear climatic shifts, particularly aridification that are seemingly so clear in Burney's core taken north of Toliara, or the ecological changes associated with shifts in the elevational position of montane flora through time are not readily apparent in the Lake Tritrivakely samples. Thus, in short it is not possible to invoke a drying up of wetland environments on the Central High Plateau during the Holocene to explain the disappearance of numerous species of waterbirds from the area. This may have happened, but current paleoecological data are insufficient to address this point.
Implications for modern conservation on Madagascar
On the basis of subfossil evidence it is clear that numerous species of birds have gone extinct on Madagascar since the start of the Holocene. The large terrestrial elephant birds, several species of large raptors, and numerous waterbirds have gone extinct. Further, several species of waterfowl that are currently extant on the island once had a much broader distribution. The last refuge for many of these waterbirds is the northwestern portion of Madagascar, a region that still has several areas of largely undisturbed aquatic ecosystems.
Within the reserve system of Madagascar there are few protected areas with extensive aquatic habitat. The only reserve named to date with freshwater habitat is the Réserve Naturelle Intégrale d'Ankarafantsika. The lakes near Antsalova, which hold important populations of several vulnerable or endangered waterbirds such as Anas bernieri, are prime candidates for protection. Furthermore, with the intense human population growth on the island and the ever expanding need to convert wetlands into rice paddy, the remaining wetlands will come under ever increasing human pressure.
ACKNOWLEDGEMENTS
The following colleagues allowed access to or lent specimen material under their care: G. Barrowclough, American Museum of Natural History, New York; R. Prys-Jones, British Museum of Natural History, Tring; C. Lefèvre, Laboratoire d'Anatomié Comparée, J.-F. Voisin and J. Cuisin, Laboratoire de Zoologie, and D. Goujet and H. Lelièvre, Laboratoire de Paléontologie, Muséum National d'Histoire Naturelle, Paris; L. Bennun, National Museums of Kenya, Nairobi; F. C. Sibley, Peabody Museum of Natural History; J. Barlow, Royal Ontario Museum, Toronto; B. Rakotosamimanana, Service de Paléontologie, Université d'Antananarivo, Antananarivo; R. Prum, University of Kansas, Museum of Natural History, Lawrence; R. Payne and R. Storer, University of Michigan Museum of Zoology, Ann Arbor; and S. Olson, National Museum of Natural History, Washington, D.C. L. Rakotozafy helped with the determination of bird subfossils held in the collections of the Université d'Antananarivo. This work was supported by grants from Conservation International, the Field Museum of Natural History, World Wide Fund for Nature, and the John D. and Catherine T. MacArthur Foundation.
REFERENCES
Andrews, C.W. 1897. On some fossil remains of carinate birds from central Madagascar. Ibis 3 (seventh series): 343-359.
Barthère, F.-M. 1915. Observations sur une Hache en os, provenant des fouilles exécutées par l'Académie Malgache, à Ampasambazimba en 1908 (Madagascar). Bulletin de la Société Préhistorique Française 12: 358-361.
Brown, L.H., Urban, E.K. & Newman, K. 1982. The birds of Africa. Vol. 1; London; Academic Press.
Burleigh, R. & Arnold, E.N. 1986. Age and dietary differences of recently extinct Indian Ocean tortoises (Geochelone s. lat.) revealed by carbon isotope analysis. Proceedings of the Royal Society of London, B 227: 137-144.
Burney, D.A. 1987a. Late Holocene vegetational change in central Madagascar. Quaternary Research 28: 130-143.
Burney, D.A. 1987b. Pre-settlement vegetation changes at Lake Tritrivakely, Madagascar. Palaeoecology of Africa 18: 357-381.
Burney, D.A. 1993. Late Holocene environmental changes in arid southwestern Madagascar. Quaternary Research 40: 98-106.
Burney, D.A. 1997. Theories and facts regarding Holocene environmental change before and after human colonization. In: Goodman, S. M. & Patterson, B. D. (eds) Natural change and human impact in Madagascar. Washington, D.C.; Smithsonian Institution Press: 75-89.
Burney, D.A., James, H.F., Grady, F.V., Rafamantanantsoa, J.-G., Ramilisonina, Wright, H.T. & Cowart, J.B. 1997. Environmental change, extinction and human activity: evidence from caves in NW Madagascar. Journal of Biogeography 24: 755-767.
Cheke, A.S. 1987. An ecological history of the Mascarene Islands, with particular reference to extinctions and introduction of land vertebrates. In: Diamond, A. W. (ed) Studies of Mascarene Island birds. Cambridge; Cambridge University Press: 5-89.
Dewar, R.E. 1997. Were people responsible for the extinction of Madagascar's subfossils, and how will we ever know? In: Goodman, S. M. & Patterson, B. D. (eds) Natural change and human impact in Madagascar. Washington, D.C.; Smithsonian Institution Press: 364-377.
Dewar, R.E. & Wright, H.T. 1993. The culture history of Madagascar. Journal of World Prehistory 7: 417-466.
Flacourt, E. de. 1658 [reprinted 1995]. Histoire de la Grande Isle de Madagascar. Edition présentée and annotée par Claude Allibert. Paris; INALCO-Karthala.
Fontoynont, M. 1909. Les gisements fossilifères d'Ampasambazimba. Bulletin de l'Académie Malgache 6: 3-8.
Gasse, F., et al. 1994. A 36 ka environmental record in the southern tropics: Lake Tritrivakely (Madagascar). Compte Rendus Académie des Sciences Paris, série II, 318: 1513-1519.
Godfrey, L.R., Jungers, W.L., Reed, K.E., Simons, E.L. & Chatrath, P.S. 1997. Subfossil lemurs: Inferences about past and present primate communities in Madagascar. In: Goodman, S. M. & Patterson, B. D. (eds) Natural change and human impact in Madagascar. Washington, D.C.; Smithsonian Institution Press: 218-256.
Goodman, S.M. 1994. Description of a new species of subfossil eagle from Madagascar: Stephanoaetus (Aves: Falconiformes) from the deposits of Ampasambazimba. Proceedings of the Biological Society of Washington 107: 421-428.
Goodman, S.M. 1996. Description of a new species of subfossil lapwing (Aves, Charadriiformes, Charadriidae, Vanellinae) from Madagascar. Bulletin du Muséum National d'Histoire Naturelle, Paris, série 4, Section C, 18: 607-614.
Goodman, S.M. in press. A description of a new species of Brachypteracias (Family Brachypteraciidae) from the Holocene of Madagascar. Ostrich.
Goodman, S.M., Langrand, O. & Razafimahaimodison, J.-C. 1994. Les oiseaux. In: Goodman, S.M. & Langrand, O. (eds) Inventaire Biologique, Forêt de Zombitse. Recherches pour le Développement, Série Sciences Biologiques, Centre d'Information et de Documentation Scientifique et Technique, Antananarivo, No. Spécial: 73-84.
Goodman, S.M. & Rakotondravony, D. 1996. The Holocene distribution of Hypogeomys (Rodentia: Muridae: Nesomyinae) on Madagascar. In: Lourenço, W. R. (ed.) Biogéographie de Madagascar. ORSTOM, Paris: 283-293.
Goodman, S.M. & Rakotozafy, L.M.A. 1995. Evidence for the existence of two species of Aquila on Madagascar during the Quaternary. Geobios 28: 241-246.
Goodman, S.M. & Rakotozafy, L.M.A. 1997. Subfossil birds from coastal sites in western and southwestern Madagascar: A paleoenvironmental reconstruction. In: Goodman, S. M. & Patterson, B. D. (eds) Natural change and human impact in Madagascar. Washington, D.C.; Smithsonian Institution Press: 257-279.
Goodman, S.M. & Ravoavy, F. 1993. Identification of bird subfossils from cave surface deposits at Anjohibe, Madagascar, with a description of a new giant Coua (Cuculidae: Couinae). Proceedings of the Biological Society of Washington 106: 24-33.
Johnsgard, P.A. 1981. The plovers, sandpipers, and snipes of the world. Lincoln; University of Nebraska Press.
Jully, A. 1898. Les tourbières d'Antsirabé et les animaux disparus de Madagascar. Notes, Reconnaissances et Explorations 1898: 1175-1183.
Lamberton, C. 1934. Contribution à la connaissance de la Faune subfossile de Madagascar. Lémuriens et Ratites. Mémoires de l'Académie Malgache 17: 1-168.
Langrand, O. 1995. Guide des oiseaux de Madagascar. Lausannae and Paris; Delachaux et Niestlé.
Langrand, O. & Goodman, S.M. 1997. Les oiseaux. In: Langrand, O. & Goodman, S. M. (eds) Inventaire Biologique, Forêt de Vohibasia et d'Isoky-Vohimena. Recherches pour le Développement, Série Sciences Biologiques, Centre d'Information et de Documentation Scientifique et Technique, Antananarivo No. 12: 131-143.
Langrand, O. & Wilmé, L. 1993. Protection des zones humides et conservation des espèces d'oiseaux endémiques de Madagascar. Proceedings of the Eighth Pan-African Ornithological Congress: 201-208.
LeNoble, A. 1940. Études sur la Géologie de Madagascar. Note 3: Le massif volcanique de l'Itasy. Mémoires de l'Académie Malgache 32: 43-80.
Lewis, A. & Pomeroy, D. 1989. A bird atlas of Kenya. Rotterdam; A. A. Balkema.
MacPhee, R.D.E. 1986. Environment, extinction, and Holocene vertebrate localities in southern Madagascar. National Geographic Research 2: 441-455.
MacPhee, R.D.E. & Burney, D.A. 1991. Dating of modified femora of extinct dwarf Hippopotamus from southern Madagascar: implications for constraining human colonization and vertebrate extinction events. Journal of Archaeological Sciences 18: 695-706.
MacPhee, R.D.E., Burney, D.A. & Wells, N.A. . 1985. Early Holocene chronology and environment of Ampasambazimba, a Malagasy subfossil lemur site. International Journal of Primatology 6: 463-489.
Mahé, J. & Sourdat, M. 1972. Sur l'extinction des Vertébrés subfossiles et l'aridification du climat dans le Sud-Ouest de Madagascar. Bulletin de la Société Géologique de France 14: 295-309.
Nicoll, M. & Langrand, O. 1989. Madagascar: Revue de la conservation et des aires protégées. Gland; World Wide Fund for Nature.
Rakotozafy, L.M.A. 1993. Étude sur des Anatidae subfossiles et leur paléoenvironnement dans les Hauts-Plateaux Malgaches. Mémoire, D.E.A., Faculté des Sciences, Université d'Antananarivo, Antananarivo, Madagascar.
Safford, R.J. 1993. The Madagascar Teal Anas bernieri: a preliminary survey from Antsalova to Morondava. Dodo 29: 95-102.
Standing, H.F. 1908. On recently discovered subfossil primates from Madagascar. Transactions of the Zoological Society of London 18: 59-162.
Stuenes, S. 1989. Taxonomy, habits, and relationships of the subfossil Madagascan hippopotami Hippopotamus lemerlei and H. madagascariensis. Journal of Vertebrate Paleontology 9: 241-268.
Tattersall, I. 1973. A note on the age of the subfossil site of Ampasambazimba, Miarinarivo Province, Malagasy Republic. American Museum Novitates 2520: 1-6.
Wetterstrom, W. & Wright, H.T. 1992. Une contribution à la paléoethnobotanique du plateau central de Madagascar. Taloha 11: 147-166.
White, E.I. 1930. Fossil hunting in Madagascar. Natural History Magazine 2 (15): 209-235.
Wiman, C. 1935. Über Aepyornithes. Nova Acta Regiae Societatis Scientiarum Upsaliensis, series 4, 9, no. 12.
Wright, H.T., Andrianaivoarivony, R., Bailiff, I., Burney, D., Haas, H., Raharijaona, V., Rakotovololona, S., Rasamuel, D. & Dewar, R. 1992. Datation absoule de sites archéologiques du centre de Madagascar - présentation des déterminations. Taloha 11: 121-145.
Table 1. Radiocarbon dates available from the three subfossil sites discussed in text
Table 2. Species of birds recovered from three subfossil sites on Madagascar. Upper case 'X' marks indicate that the species still occurs in the general region of the subfossil site.
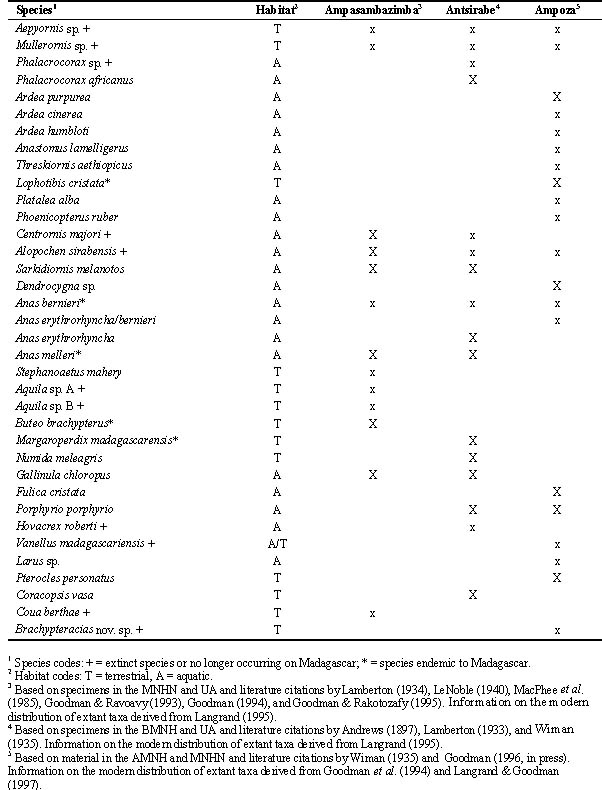
Table 3. Summary information on various aspects of the avifauna of the three subfossil sites.
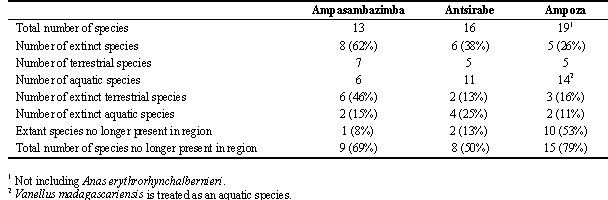
Fig. 1. Map of Madagascar showing subfossil sites mentioned in the text. The information on modern habitats is derived from Nicoll & Langrand (1989).